
Climate change is an important factor affecting species dispersal and distribution through time and the accelerated global warming has currently concerned decision makers and conservationists. Because protected areas are spatially static, species extinction risk is generally expected to increase under climate change scenarios as a consequence of range shift and decrease. This study aims to understand the current conservation status of Actinote quadra, a neotropical threatened butterfly species, as well as predict how it will be in the future. By coupling ecological niche modeling and climatic simulations, we predicted the species distribution in different future global warming scenarios (2050 and 2070) and estimated the proportion of species range overlapping protected areas through time. Our findings showed a generalized decrease of the potential distribution of A. quadra in the end of 21st century, with the most prominent range loss predicted to occur out of protected areas. Although climate change will potentially drive A. quadra into reserves, the predicted range collapse would be enough to increase its extinction risk from vulnerable, like currently categorized, to the status of critically endangered in accordance to IUCN red list criteria. Taking into account the fragmented and discontinuous landscapes across the Atlantic Forest's hotspot, we propose a conservation strategy for A. quadra based on potential ecological corridors linking climatically suitable areas and discuss the need for amplifying and connecting the current protected areas to maintain this threatened species at longer time under a global warming scenario.
Creating endangered species lists and establishing protected areas are traditional actions taken by conservationists and decision makers and have long been implemented for conservation policies worldwide (see Colyvan et al., 1999; IUCN, 2001). Recently, some practical approaches as reserve-selection methods seek to maximize the current biodiversity within protected areas (Margules and Pressey, 2000). Such protected areas are relevant to preserve species suffering human pressures in a given period, especially within hotspots of biodiversity (Myers et al., 2000). However, the protected areas preserve species in a static space while species geographical ranges are dynamics through time (Badgley et al., 2008). Changes in environmental conditions, for instance, might considerably reduce the species geographical range or drive it away from its current locations by tracking environmentally suitable areas through time (“habitat tracking” sensu Eldredge et al., 2005). As a consequence, the persistence of viable populations within protected areas could be affected under environmental change scenarios, like the current global warming, and decrease the effectiveness of conservation practices in the future (Araújo et al., 2004).
Recently, different modeling procedures have been proposed to infer species dynamics under future climatic scenarios. For instance, coupling ecological niche models (ENM) and climatic simulations have been extensively used to predict the potential distribution of species and assess the effectiveness of current protected areas in maintain biodiversity in a world of future changes (see examples in Araújo et al., 2004; Loyola et al., 2012; Prieto-Torres et al., 2015). ENMs predictions are especially important to understand the distributional dynamics of species under climate change and to delineate conservation strategies more efficient to keep these threatened species preserved at longer time in the future.
In Brazil, the threatened species list (Brazilian red-list) has currently 130 terrestrial invertebrate species in different categories of threat (Machado et al., 2008, similar to the proposed by IUCN, 2001), among them, 55 butterfly species (Freitas and Marini-Filho, 2011). Butterflies represent around a half of threatened Brazilian terrestrial invertebrates species and have been considered important for conservation issues because they are a key-group to maintain relevant ecosystem services like pollination (Freitas et al., 2006). Pollination, for instance, is crucial to support native populations of wildflowers, as well as for keeping economically viable some human activities like agriculture (Melathopoulos et al., 2015). Moreover, the most species of threatened butterflies occur in the Atlantic Forest domain (Freitas and Marini-Filho, 2011), a Neotropical hotspot of biodiversity (Myers et al., 2000). Thus, conservation studies generating new knowledge about threatened butterflies in Atlantic Forest region are important for conservation actions in the future.
In 2011 the ICMBio, an Institution from Brazilian Environmental Ministry, published the Action Plan for conservation of threatened Lepidoptera species (Freitas and Marini-Filho, 2011) in which Actinote quadra (Schaus 1902) (Lepidoptera: Nymphalidae) was listed as vulnerable. This species is endemic to mountainous environments (mainly>1000m above sea level, only 3 occurrence data below 1000m) of Atlantic Forest (Gomes et al., 2014), across states of São Paulo, Minas Gerais and Rio de Janeiro, and present populations fragmented and restricted across less than 2.000km2 (Freitas et al., 2009). According to the ICMBio action plan, a plausible conservation strategy for A. quadra is search for new viable populations and maintain intact its occurrence areas; i.e. ensuring that populations will not decrease in the future (Freitas and Marini-Filho, 2011).
Here, we set the potential conservation status of A. quadra under the global warming scenario in the future by coupling ecological niche modeling and climatic simulations. We specifically assessed the effectiveness of current protected areas in maintain climatically suitable areas for A. quadra during the 21st century, and aimed to answer three questions: What is the current state of conservation of A. quadra and how it will be in the future? Will the climatically suitable areas reduce in the future? Are the current protected areas efficient to keep A. quadra across climatically suitable areas in the future? These questions are important for understanding the dynamics of potential impacts that A. quadra will suffer under the future global warming, developing thus more efficient conservation strategies in the future. Although conservation strategies for unique species often have not been important in political decisions, the modeling approach implemented here has been a helpful way (or maybe the unique scientifically reliable way) to predict potential distributional dynamics and infer extinction risk of species in the future. As a consequence, predicting the impact of global warming on unique species is a first important step for improving concepts and tools to implement conservation protocols in a dynamical world rather than in a static context, strengthening the conservation practices under global change scenarios, and developing more efficient conservation strategies for longer time by further focusing on the entire biodiversity.
Material and methodsEcological niche modelingTo predict the current and the future potential distribution of A. quadra, we used techniques of ecological niche modeling (ENMs). Niche modeling basically estimates associations between environmental aspects (most often climate) and known species occurrences to characterize the range of conditions under which the species’ populations survive, the called environmental (or climatic) suitability (Franklin, 2009; Peterson et al., 2011). A total of 16 occurrence records of A. quadra were obtained in Gomes et al. (2014) and mapped in a grid of cells with 2.5′ resolution (∼4.5km×4.5km at the equator). We used this resolution because the occurrence points are practically unique by cell (only unique duplicated point was removed) (Table S1) and the climate layers are available for different temporal scenarios. All 15 occurrence records were used for calibration of distribution modeling.
To predict the current and the future potential distribution of A. quadra, we used techniques of ecological niche modeling (ENMs). Niche modeling basically estimates associations between environmental aspects (most often climate) and known species occurrences to characterize the range of conditions under which the species’ populations survive, the called environmental (or climatic) suitability (Franklin, 2009; Peterson et al., 2011). A total of 16 occurrence records of A. quadra were obtained in Gomes et al. (2014) and mapped in a grid of cells with 2.5′ resolution (∼4.5km×4.5km at the equator). We used this resolution because the occurrence points are practically unique by cell (only unique duplicated point was removed) (Table S1) and the climate layers are available for different temporal scenarios. All 15 occurrence records were used for calibration of distribution modeling.
To characterize the environmental aspects of Neotropical region, we used five bioclimatic variables from WorldClim database (www.worldclim.org): Annual Mean Temperature, Mean Diurnal Range, Isothermality, Precipitation of Wettest Quarter and Precipitation of Driest Quarter. These five bioclimatic variables present low multicollinearity and were selected by factorial analysis with Varimax rotation from 19 bioclimatic variables (www.worldclim.org/bioclim). The climate predictors present 2.5 arc-resolution (∼4.5km×4.5km at the equator) and were built for current and future scenarios. Future scenario was represented by climatic conditions at 2050 and 2070 from two Atmosphere-Ocean Circular Models (AOGCMs) – CCSM and MIROC – using the carbon emission scenario RCP45.
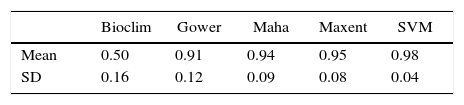
Because predictions differ among ENM methods (see Barry and Elith, 2006; Diniz-Filho et al., 2009), we used five algorithms based on different modeling methods and assumptions for inferring A. quadra's potential distribution: three simple methods based only on presence records, (1) Bioclim (Nix, 1986), (2) Mahalanobis distance (Farber and Kadmon, 2003), (3) Domain (Gower distance; Carpenter et al., 1993); and two machine-learning methods based on presence-background records, (4) Support vector machines (SVM) (Tax and Duin, 2004), (5) Maximum Entropy – Maxent (Phillips and Dudik, 2008). The distribution of A. quadra was first modeled for current climate and then projected onto the future conditions at 2050 and 2070. All algorithms were run in the R-package “dismo” (Hijmans et al., 2015) using specific functions matching each algorithm, except SVM (run using the function ksvm from R-package “kernlab” (Karatzoglou et al., 2004). The Bioclim from R-package dismo computes the suitability of a location by comparing the values of environmental variables at any location to a percentile distribution of the values at known locations of occurrence (“training data”, the closer to the 50th percentile – the median, the more suitable the location is) instead of predicts binary presences and absences from traditional rectangular envelopes (see Hijmans et al., 2015). Mahalanobis distance and Domain compute the environmental distance from each grid cell to species ecological optimum (established from its occurrence records; Carpenter et al., 1993; Farber and Kadmon, 2003). Mahalanobis distance uses the multivariate mean and covariance structure among the climatic variables linked to occurrence records for establishing the center of the bioclimatic envelope (the climatic optimum), and then calculates distances to this centroid as a proxy for climatic suitability. On the other hand, Domain computes the Gower distance, in a given climatic space, between each grid cell and the closest point related to an occurrence record. Thus, Domain does not consider a unique climatic optimum (centroid) for the focal species, whereas Mahalanobis distance establishes the climatic optimum based on a multivariate space. The machine-learning methods were fitted using the simplest configurations for classification tasks, like linear kernel function and c-SVC type for SVM (with probabilistic output) and only the linear feature to combine climatic predictors in Maxent (with logistic output). Because A. quadra has a few occurrence points and we aim obtain its potential distribution, simple modeling configurations are recommended (see Varela et al., 2011). All other parameters from SVM (e.g., cost parameter – C) and Maxent (e.g., beta, gamma, and prevalence; see Elith et al., 2006) were used as in default.
Background points were randomly selected throughout the Neotropical grid cells (excepting cells with presences) keeping species prevalence equal to 0.5. This approach was based on recent studies suggesting that the extent of the geographical region in which the background points are taken have important influences for prediction and performance of ENMs (e.g., Thuiller et al., 2004; Van der Wal et al., 2009). Although A. quadra is currently an endemic species from Atlantic Forest, we used the entire Neotropical region to select background points because there is no reason expected to prevent species dispersal throughout Neotropics (i.e., we considered the entire Neotropical region as historically accessible for A. quadra, see Barve et al., 2011). Thus, selecting background points throughout the Neotropical region essentially represents a compromise between generating models that neither underpredict (i.e., generalist models) nor overpredict (i.e., precise models) the species distribution (Van der Wal et al., 2009). Because this wide background throughout the Neotropics may be considered unrealistic and misclassified as historically accessible area for the focal species, we performed sensitivity analyses by building the ENMs from two additional backgrounds based strictly on the restricted conditions in which A. quadra occur nowadays: (1) taking into account that focal species is currently restricted to Atlantic Forest across Southern Brazil, we considered the 23°S zone from UTM system as the first additional background (regional background); and (2) considering that A. quadra is a mountainous species currently restricted to high altitudes, we restricted that former background (23°S-UTM zone) to areas higher than 700m above sea level (altitudinal background). Background points were used to calibrate the ENMs based on presence-background observations (SVM and Maxent) and compute confusion matrix to evaluate model performance (see below).
For each algorithm, 20 models were initially fitted using a two-fold partitioning criterion (75% and 25%) for training models and evaluating their performance. The 75–25% of training and testing data were randomly selected for each one of 20 initial models. These repetitions are expected to minimize the spatial structure between the training and testing data sets, providing less biased evaluations. For evaluation, we used the True Skilled Statistic (TSS) whose values range from −1 to 1; negative or near zero values indicate that model predictions are not different from a randomly generated model, whereas models with values closer to 1 are said to be excellent. In general, acceptable models have TSS values above 0.5 (Allouche et al., 2006).
Next, the lowest suitability value from an occurrence record was used as cut-off threshold (LPT – lowest presence threshold) to compute predicted binary maps (Jiménez-Valverde et al., 2011). By following the ensemble approach (Araújo and New, 2007), the 20 binary maps belonging a same algorithm were then concatenated to compute the frequency of presences predicted for each grid cell, yielding a single consensus map per algorithm and climatic scenario (present, 2050 and 2070). Similarly, we obtained the final consensus map for each climatic scenario by computing frequencies from all algorithms.
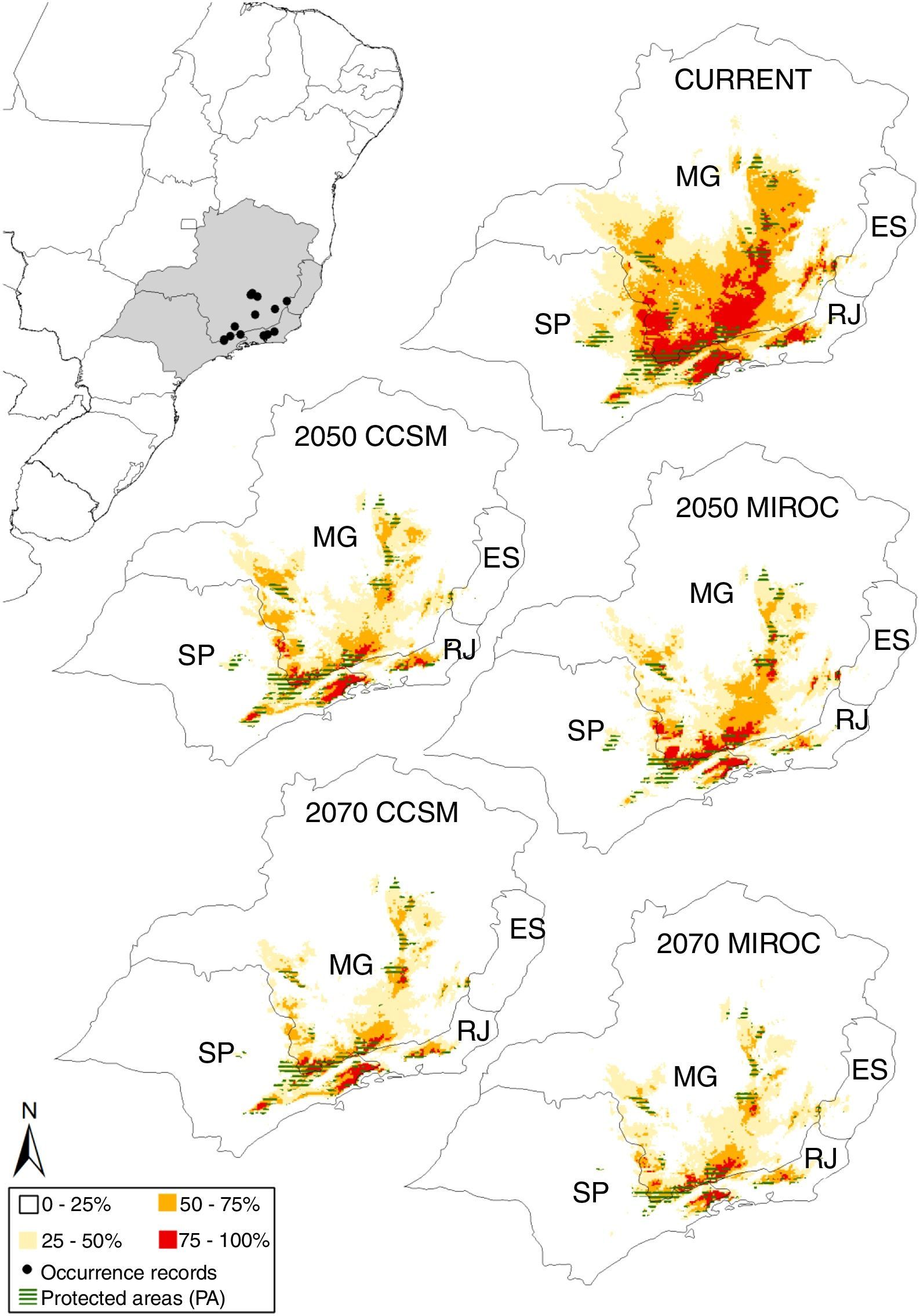
The potential distribution of A. quadra was classified in quartiles of predicted frequencies (0–25%, 25–50%, 50–75%, and 75–100% consensual classes) for present, 2050 and 2070. This less permissive quartile gradient of frequencies permit us to analyze the extinction risk and climate change effects across specific regions where A. quadra was more (quartile 75–100%) or less (quartile 0–25%) reliably predicted to occur through time. Finally, we observed that the potential distribution of A. quadra was predicted to be restricted to the southeastern Brazil (none model predicted A. quadra to potentially occur out of this region). We cut the quartile frequency map of each scenario using the 23°S zone from UTM system and, therefore, results are presented only for this extent, even ENMs have been built using Neotropical background.
Analyzing extinction risk and climate change effectsWe mapped and quantified the climatically suitable area (km2) of A. quadra for each quartile from all climatic scenarios (current, 2050 and 2070) and used the classification schemes from IUCN red list criteria (http://www.iucnredlist.org/) to evaluate its extinction risk through time. IUCN criteria for threat classification rely on many ecological aspects such as population dynamic and size, extent of occurrence and/or area of occupancy, and quantitative analyses (population viability). In our case, because ENMs yield potential distributions (and not realized ones; see discussion in Jiménez-Valverde et al., 2008), the IUCN absolute cutoffs (e.g., extent of occurrence, in km2 – criteria B – or population size, in number of individuals – criteria C and D) do not represent a fine estimate for extinction risk from niche modeling. For our purpose, we believe that more plausible criteria should be based on relative cutoffs, (decrease/increase, in percentage) through time. However, the IUCN criteria based on relative cutoffs are directly based on population decrease rather than geographical range dynamics, given by criterion A. Thus, we linked population dynamics with our estimates of potential distribution by assuming a given fixed population density of A. quadra and used the variance values established by IUCN criteria A4b,c to determine the species extinction risk through time based on range dynamics. For example, A. quadra will be Critically Endangered (CR) at the end of 21st century if impacts from climate changes yield in a range decrease ≥80%, Endangered (EN) if ≥50%, Vulnerable (VU) if ≥30%, and unthreatened (Near Threatened – NT – or Least Concern – LC) in other cases. This approach indicates how much the global warming will affect the extinction risk of A. quadra by constrain their populations at the end of 21st century as a consequence of decreases in its predicted geographical range.
Finally, we computed the area of potential distribution (km2) of A. quadra into protected areas (PA) for three consensual quartiles (25–50%, 51–75% and 76–100%) and analyzed the proportion of PA in relation to the total species distribution area (TA, total area) for each climatic scenario (current, 2050 and 2070). Protected areas were obtained from Brazilian Environmental Ministry (http://www.mma.gov.br) and cut for the Southern Brazil extension (UTM 23S projection). We considered all classifications of protected areas (Ecological Station, Biological Reserve, National Park, Natural Monument, Wildlife Refuge, Environmental Protection Area, Relevant Ecological Interest Area, National Forest, Extractive Reserve, Faunal Reserve, Sustainable Developing Reserve and Private National Heritage Reserve) in federal, state, and municipality political domains.
ResultsThe ecological niche models provided reliable predictions from all algorithms since TSS values were greater than 0.5 (Table 1), except for some Bioclim’ predictions. Niche models predicted the potential distribution of A. quadra in present-day across the states of São Paulo, Minas Gerais and Rio de Janeiro in Southeastern Brazil, regardless of used background (Fig. 1, S1). Because we consider those restricted background by unrealistically constraining the geographical range and dispersal opportunity of A. quadra throughout past climate changes like Quaternary glaciations, we will present just the result from Neotropical background (Fig. 1).
 overlapping extant protected areas.")
The ecological niche models provided reliable predictions from all algorithms since TSS values were greater than 0.5 (Table 1), except for some Bioclim’ predictions. Niche models predicted the potential distribution of A. quadra in present-day across the states of São Paulo, Minas Gerais and Rio de Janeiro in Southeastern Brazil, regardless of used background (Fig. 1, S1). Because we consider those restricted background by unrealistically constraining the geographical range and dispersal opportunity of A. quadra throughout past climate changes like Quaternary glaciations, we will present just the result from Neotropical background (Fig. 1).
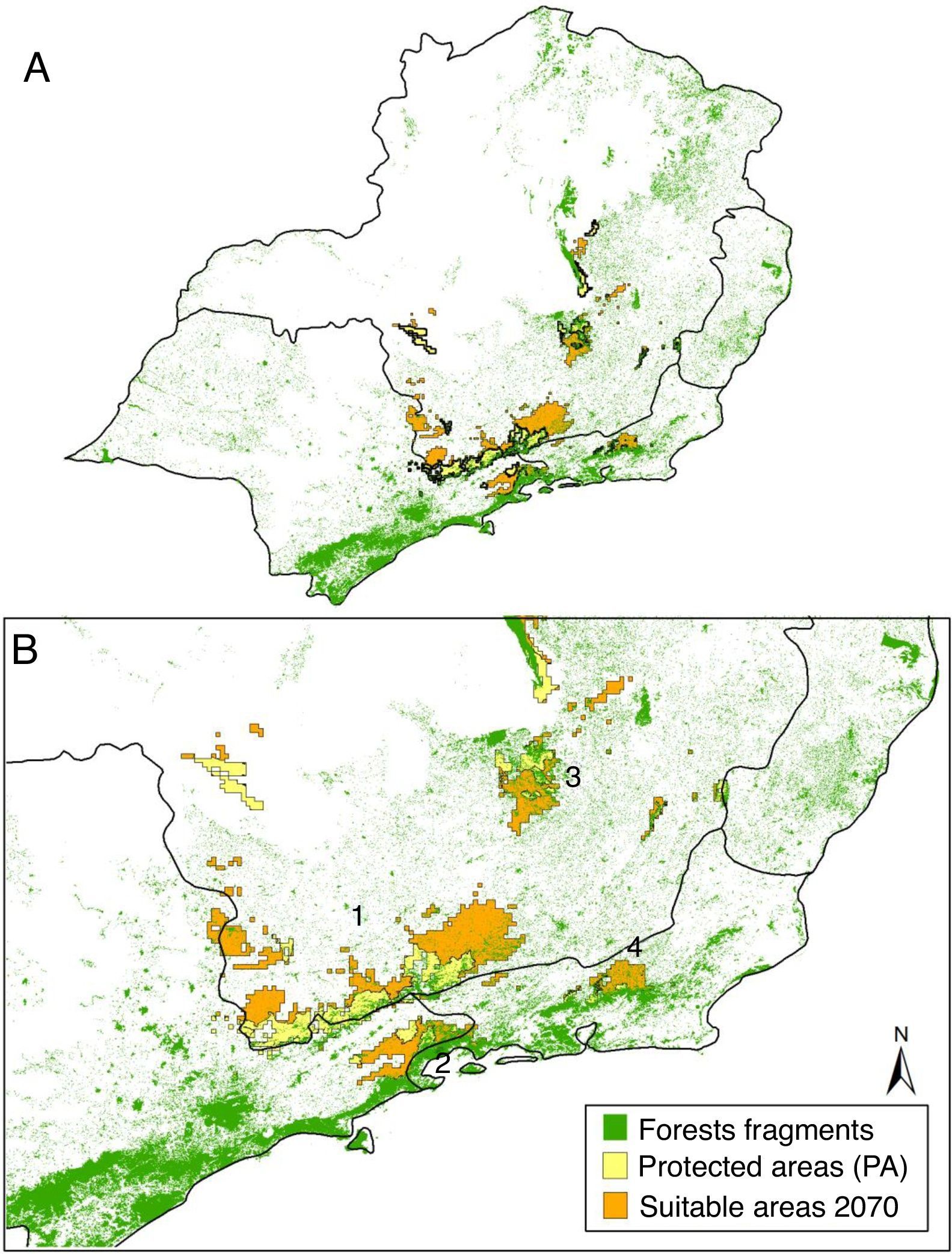
Predictions indicate four disconnected mountain blocks as climatically suitable for the species across Atlantic Forest, the Serra do Mar, the South Mantiqueira, and the mountain blocks in Rio de Janeiro and Minas Gerais (Fig. 2), like expected from field observations (Gomes et al., 2014).

Potential distribution of A. quadra matching four historically isolated mountain blocks: 1 – South Mantiqueira block, 2 – Serra do Mar block, 3 – Minas Gerais mountains block, and 4 – Rio de Janeiro mountains block. Yellow and orange areas represent the current potential distribution of A. quadra within PAs and its suitable areas on 2070 scenario. Green fragments indicate the remnants of Atlantic Forest.
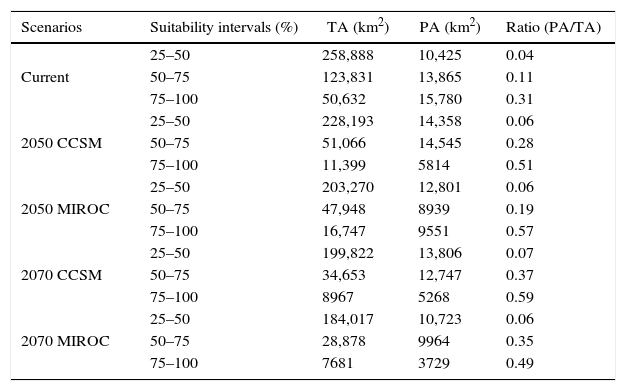
The species potential distribution (total area: TA) decreases under the future conditions regardless of climatic scenario (CCSM or MIROC), remaining only 15% of the most suitable original area in 2070 (i.e., a range loss around 85% at the end of 21st century; Table 2). Similarly, the potential distribution overlapping protected areas (PA) also will decrease in the future, although in less severe levels (remaining 25% of the protected original area).
The total distribution area (TA) and the protected area (PA) in current, 2050 and 2070 climatic scenarios for three quartiles of suitability.
| Scenarios | Suitability intervals (%) | TA (km2) | PA (km2) | Ratio (PA/TA) |
|---|---|---|---|---|
| Current | 25–50 | 258,888 | 10,425 | 0.04 |
| 50–75 | 123,831 | 13,865 | 0.11 | |
| 75–100 | 50,632 | 15,780 | 0.31 | |
| 2050 CCSM | 25–50 | 228,193 | 14,358 | 0.06 |
| 50–75 | 51,066 | 14,545 | 0.28 | |
| 75–100 | 11,399 | 5814 | 0.51 | |
| 2050 MIROC | 25–50 | 203,270 | 12,801 | 0.06 |
| 50–75 | 47,948 | 8939 | 0.19 | |
| 75–100 | 16,747 | 9551 | 0.57 | |
| 2070 CCSM | 25–50 | 199,822 | 13,806 | 0.07 |
| 50–75 | 34,653 | 12,747 | 0.37 | |
| 75–100 | 8967 | 5268 | 0.59 | |
| 2070 MIROC | 25–50 | 184,017 | 10,723 | 0.06 |
| 50–75 | 28,878 | 9964 | 0.35 | |
| 75–100 | 7681 | 3729 | 0.49 |
The ratio PA/TA continuously increases from present toward 2050 and 2070, indicating that habitat loss will be higher outside protected areas than within them. Such pattern occurs for all consensus levels, but it is more evident for the most suitable areas (frequency of 75–100%; Table 2).
DiscussionOur findings highlight potential effects of climate warming upon distributional dynamics of A. quadra. A reduction around 85% of species potential distribution was predicted for the end of 21st century. This scenario is expected because A. quadra is endemic to mountainous (>800m above sea level) environments of Atlantic Forest with mean temperature often below 19°C (Freitas et al., 2009; Gomes et al., 2014). With increasing temperatures in the future, these climate conditions will occur on even higher altitudes or may even no longer exist in lower mountains. Anyway, it is expected that mountainous species like A. quadra will be increasingly restricted to the highest areas of the Southeastern Brazil in response to global warming (see debate of species characteristic and its distributional dynamics in Carnaval et al., 2014).
Along with an expressive range reduction, our findings showed that the greatest range loss will especially occur out of protected areas. In a first glance, current protected areas would be efficient to maintain viable population of A. quadra across climatically suitable areas in the future. However, these protected areas represent a small part of suitable area for A. quadra in 2050 and 2070. Based on IUCN (2001) criteria, the extinction risk of A. quadra will increase even being preserved in some climatically suitable locations. With a predicted range reduction around 85%, the species will become critically endangered at the end of 21st century instead of vulnerable as it is considered nowadays. If we consider that A. quadra will survive at the end of 21st century just across climatically suitable areas within extant protected areas because of continuous habitat loss outside them, the range reduction is still higher (92%; i.e., comparing the total suitable area in the present with the protected suitable area in the future) and extinction risk would further increase.
Therefore, our models predict A. quadra will substantially be more prone to extinction in the future than today due to expressive range reduction, and extinction risk will increase especially out of reserves. Based on these results, we conclude that extant protected areas will not be enough or efficient to maintain A. quadra in the future, which signalize the need for improving conservation strategies for A. quadra for longer time.
There are two complementary strategies well established to improve conservation actions in a biogeographical scale: (1) create new reserves to increase the protected area, and (2) create ecological corridors linking extant protected areas (Whittaker et al., 2005; Ladle and Whittaker, 2011). Regardless of adopted focus, Hannah et al. (2007) reinforce that conservation strategies should be proposed basing on ecological theories. Thus, by considering isolated and fragmented protected areas as islands in a landscape-scale context, we propose here a two-steps conservation planning for A. quadra by coupling both previous focuses based on the principles of island biogeography (Spellerberg and Sawyer, 1999) and metapopulation dynamics (Hanski and Gilpin, 1991). Firstly, we propose establishing potential ecological corridors linking current protected areas. In a second instant, we propose increase the area of extant reserves or creating new reserves around extant ones instead of creating isolated reserves into anthropogenic matrix. In both steps, it is important that involved protected areas are climatically suitable in the future.
Ecological corridors are important to facilitate colonization of putative isolated areas, keep gene flow among populations from different protected areas, and decrease local extinction risk (Wilson and Willis, 1975; Simberloff and Cox, 1987). Because A. quadra occupy the top of mountains and was predicted to occur across four historically disconnected mountain blocks (the Serra do Mar, the South Mantiqueira, and the mountain blocks in Rio de Janeiro and Minas Gerais; Fig. 2), it is fundamental that corridors connect protected areas within each mountain block. On one hand, dispersal among the four historically disconnected blocks is naturally unlikely and original vegetation was practically decimated across the valleys (Fig. 2; SOS Mata Atlantica, 2014), so corridors linking mountain blocks would be ineffective to promote colonization and gene flow. On the other hand, there is around 22% of original Atlantic Forest in remnants across private properties (e.g., farms), mainly by determination of Brazilian Forest law (MMA/IBAMA, 2012). Thus, we propose that ecological corridors should be set along the private remaining Atlantic Forest preserved by Brazilian Forest law. Because these remnants of natural vegetation cannot rightfully be vanished from private properties, regardless of economical interest, we believe this aspect of ecological corridors is operationally feasible. At same time, it is expected that corridors across climatically suitable areas are also ecologically meaningful to maintain viable populations of A. quadra within protected areas in the future (Fig. 2).
Along with connections, we emphasize that it is also important increasing the extant protected areas into each mountain block matching climatically suitable areas for focal species. Although the creation of new reserves is problematic in this context of A. quadra due to generalized conflicts of interest for land use in Atlantic Forest domain, wider and suitable protected areas are fundamental to keep larger and growing populations at longer time (i.e., source populations), decreasing the populations extinction risk at local scales (Hanski and Gilpin, 1991).
The questions raised and answered in this study are important for understanding the dynamics of possible impacts that a specific species, A. quadra, will suffer under the future global warming. Although conservation strategies for unique species often have not been important in political decisions, the modeling approach implemented here based on ecological niche models has been a helpful tool (or maybe the unique scientifically reliable way) to predict potential distributional dynamics and infer extinction risk of species in the future. As a consequence, predicting the impact of global warming on unique species is a first important step for improving concepts and tools to implement conservation protocols in a dynamical world rather than in a static context, strengthening the conservation practices under global change scenarios, and developing more efficient conservation strategies for longer time by further focusing on the entire biodiversity.
Conflicts of interestThe authors declare no conflicts of interest.
The authors thank Jessica Trujillo for support about the species occurrence and two anonymous reviewers for comments. This work was partially supported by FAPEG (financial support to MLR: 2012/1026.700.1086) and CNPq (graduate scholarship to TSS). TSS thanks the PPG Ecologia Unicamp by space for working and RBF thanks Universidade Católica de Santos by logistic support.
The following are the supplementary data to this article:

