






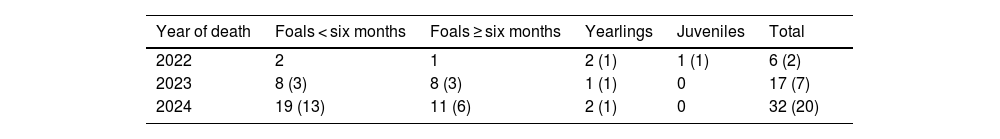
The absence of top predators can lead to significant ecological consequences, especially in association with changes in bottom-up ecosystem regulation. In the context of biological invasions, the enemy release hypothesis explains how invasive alien species can reach high densities in the absence of natural antagonists, often leading to negative effects on biodiversity and ecosystem services. Feral equids frequently reach problematic densities in natural environments, where their management faces operational and communication challenges. In this study, we evaluated the impact of puma (Puma concolor) presence on the demography of a feral horse (Equus caballus) population in an Argentinian grassland nature reserve. We compared periods with (2023–2024) and without predation (1995–2002). Foal survival was significantly lower in 2023–2024, while adult survival increased compared to 1995–2002. These changes, coupled with direct evidence of puma predation on young horse carcasses, suggest that pumas are affecting population growth rates by reducing recruitment, and may limit population size. Concomitantly, mean female body condition improved in 2023–2024, suggesting that the loss of foals to predation may reduce lactation-related energy costs. These findings suggest that pumas could play an important role in the dynamics of feral horse populations, marking the first documented case of such a regulation in South America. Incorporating predator activity into management strategies could reduce the need for direct interventions, offering a more sustainable approach to controlling the ecological impacts of feral horse populations.

Large ungulates may be regulated by bottom-up and by top-down processes (Grange and Duncan, 2006). The first are linked to ecosystem primary production, carrying capacity (K) and density‐dependence (Bonenfant et al., 2009). Top-down regulation, in turn, depends on predator abundance and activity, which can affect population dynamics and behaviour (Skogland, 1991). The debate over which of these forces dominates ecosystem regulation seems to be a thing of the past since both are nowadays regarded as playing a role to some extent in this dynamic (Lemmens et al., 2018; Pierce et al., 2012). In particular several articles highlight the role of top predators in ecosystems and trophic cascades, and the consequences of their absence (Burgos et al., 2024; Estes et al., 2011; Morris and Letnic, 2017; Ritchie et al., 2012).
The role of predators as potential agents of regulation of prey populations is of particular interest in the context of biological invasions. The enemy release hypothesis has been proposed as an explanation for the success of certain species that reach high abundances in places where they are introduced, after leaving behind their natural antagonists in their native area of distribution (Blossey, 2011), and is one of the most cited explanations of invasion success among specialists in the field (Enders et al., 2018; Liu and Stiling, 2006). To what extent can the absence of a predator facilitate invasion? And, in addition, could a predator limit the invasion capacity of an introduced prey? Despite the theoretical and applied interest of these questions, empirical evidence in this regard is very scarce, particularly in the case of interactions between vertebrates (Brian and Catford, 2023; Heger and Jeschke, 2018; Twining et al., 2022). The effect of predation will depend on different features, including interactions between multiple predators (Garneau et al., 2007), human disturbance (Guerisoli et al., 2019; Petridou et al., 2023), differences in the behaviour and needs of each sex of the carnivore involved (Cristescu et al., 2022), and on the behaviour and life cycle of prey species (Bates-Mundell et al., 2024; Harmsen et al., 2011). This could result in variations in spatio-temporal patterns in predation events and so, in their effects on the prey population. On the other hand, the concentration of predation in certain life stages, particularly on young, could have secondary effects on the population by reducing intraspecific competition or by freeing mothers from the energy costs of feeding their offspring (Berg et al., 2023; Middleton et al., 2013). The joint analysis of these components helps to adequately measure the magnitude of the eventual effect of predation on the population dynamics of the prey.
Populations of feral equids on natural and seminatural environments frequently reach high densities that result in conservation setbacks (Nuñez et al., 2016), and their management is often methodologically challenging and elicits strong emotional responses from some sectors of society (Scasta et al., 2020; Scorolli, 2018). Horses (Equus caballus) are considered an invasive alien species in Argentina, where they have been able to establish and thrive in several natural and seminatural habitats (MAyDS, 2021; Novillo and Ojeda, 2008). The most thoroughly studied population resides in Ernesto Tornquist Provincial Park (ETPP), the only state nature reserve in the country that protects the Pampean mountain grasslands. It is located in the Ventania mountain system, which include relicts of pampas grassland and represent an island of biodiversity with many endemic species (Kristensen and Frangi, 1995). These mountains also deliver valuable ecosystem services, such as water provision for neighbouring cities (Mara et al., 2020; Montico et al., 2019).
In 1942, a group of horses was introduced to ETPP. Since then, the population has increased, favoured by the almost total absence of management, with a unique removal of individuals carried out in 2006–2007 (Scorolli, 2018). Horses cause significant changes in the structure and composition of the park vegetation (de Villalobos and Zalba, 2010; Loydi et al., 2012), promote the arrival and spread of exotic woody plants (de Villalobos et al., 2011), and reduce the abundance and reproductive success of grassland birds (Zalba and Cozzani, 2004), among other ecological impacts. Demographic and body condition analyses of the ETPP feral horse population were carried out between 1995 and 2014 (Scorolli, 2021; Scorolli and Lopez Cazorla, 2010), and resumed in 2022.
The puma (Puma concolor) has been the only large carnivore present in the pampas ecosystem since the regional extinction of the jaguar (Panthera onca) more than a century ago (Agnolin et al., 2016). Puma geographic range in Argentina suffered a major contraction in the 19th Century due to direct human persecution by livestock ranchers, which led to its functional extinction in the Pampean grasslands (De Lucca, 2011). However, a recent shift from cattle ranching to agriculture that reduced direct persecution may have allowed the puma to recolonize different areas of its past distribution range, including the Ventania mountains, and also to increase its local abundance (De Lucca and Chimento, 2020).
Over the past three years, we noticed an increase in puma sightings at ETPP and detected several carcasses of juvenile horses showing signs of predation consistent with puma attacks. Most feral horse populations live in environments where there is little predation pressure (Ransom et al., 2016). However, as with other wild equids, there are documented cases where predation seems to limit their populations (Grange and Duncan, 2006; Greger and Romney, 1999; Lagos Abarzuza, 2013; Turner et al., 1992; Van Duyne et al., 2009). In this work, we compare survival rates of female feral horses across different age classes in the ETPP population between 2023 and 2024 with our previous published data from 1995 to 2002 when predation was almost absent (Scorolli and Lopez Cazorla, 2010). This comparison could help to elucidate the effect of puma predation on the demography of this population. Some studies in the western US have reported a preference of puma for foals in feral horse populations (Andreasen et al., 2021; Greger and Romney, 1999; Iacono et al., 2024; Turner and Morrison, 2001), so we expect to find a decrease in survival in the younger age classes (foals and yearlings) in the ETPP in the period with puma predation (2022–2024). In addition, we expect a smaller proportion of juveniles in the feral horse population in Tornquist Park in 2023–2024 compared to the 1995–2002 period.
We also analysed spatio-temporal patterns of puma predation events that could modulate its effects on the feral horse population. In some cases of puma predation on feral horse populations, the distribution of foal carcasses has been concentrated in areas most heavily used by the horses during spring and summer (Turner, 2015). Other authors, studying puma predation on native species such as the vicuña (Vicugna vicugna) in mountain environments, report a disproportionately high frequency of hunting in topographically complex sites (Donadio and Buskirk, 2016; Smith et al., 2019). Because predation on foals in other feral horse populations occurs mostly in summer (Andreasen et al., 2021; Greger and Romney, 1999; Turner et al., 2012), we hypothesized that the same pattern will be evident at ETPP.
Finally, as adult females subsidise foals through lactation, and this can affect their body condition (Bostal et al., 2024; Zabek et al., 2016), we propose that the loss of foals due to puma predation will reduce the mothers' energy cost of lactation, improving their body condition. The comparison of current body condition of mares at ETPP with data from periods without foal predation (2001–2002, when the population was approaching K, and 2012–2013, when it was near 50% of K, Scorolli, 2021), will allow us to evaluate whether predation on foals has a positive impact on the body condition of adult females.
We hypothesize that puma predation plays a limiting role in the feral horse population dynamics in the ETPP and should be considered a valuable component in a management strategy.
Materials and methodsStudy areaErnesto Tornquist Provincial Park (ETPP), located in southwestern Buenos Aires province (Argentina, 38° 00′–38° 07′S and 61° 52′–62° 03′W), is a protected area comprising 6700 ha of mountain grasslands, ranging in elevation from 450 to 1176 m asl. It has a temperate climate with an average annual temperature of 14 °C and average annual rainfall of 800 mm. Precipitation is concentrated in two wet seasons, in spring and autumn (Burgos, 1968). Ernesto Tornquist Provincial Park is very important for the conservation of Pampean grassland biodiversity, sheltering many endemic animal and plant species in a context of intense agricultural transformation (Guerrero and Apodaca, 2022; Kristensen and Frangi, 1995). The main vegetation comprises steppe or substeppe grasslands dominated by needlegrass (Nasella spp. and Piptochaetium spp.; Cabrera, 1976; Kristensen and Frangi, 1995). The park has areas that have been heavily invaded by woody plants, including pines, brooms, and wild cherries (Amodeo and Zalba, 2013; Brancatelli et al., 2022; Sanhueza and Zalba, 2012). At ETPP, three distinct types of grasslands are identified: lower (near piedmont at 550 m asl), medium (850 m asl), and upper (1025 m asl) (Kristensen and Frangi, 1995; Pérez and Frangi, 2000). Feral horses predominantly inhabit the lower grasslands, which offer easier access and gentler slopes compared to the steep hillsides or rocky areas found higher up. These lower areas are dotted with numerous water sources associated with streambeds, that provide a reliable year-round water source. The horse distribution pattern is particularly evident during the spring and summer (Scorolli, 1998).
Data collectionFeral horses are organized in harems with female defence polygyny (Fig. 1, Boyd et al., 2016). Between June 2022 and October 2024, each harem-band of feral horses at ETPP was observed with the aid of binoculars (10 × 50) during surveys of two consecutive days occurring once or twice a month (the latter only for October and December 2023), by traveling on foot a route that covered almost the whole 20 km2 area used by the population. Individuals were identified based on coat colour and white marks on the face and legs (Berger, 1986; Linklater et al., 2004; Scorolli and Lopez Cazorla, 2010). Horses were sexed by direct observation of genitalia and classified into four age classes based on their body size, and length of tail and mane: foals (<1 yr), yearlings (<2 yr), juveniles (<3 yr), and adults (≥3 yr). Demographic data for this population from 1995 to 2002 were taken from Scorolli (2007). To facilitate future comparisons, we divided the 1995–2002 dataset into two subperiods: 1995–1998, when horse density was lower than 30 horses/km², and 1999–2002, when it was higher. For each subperiod, we calculated the mean number of females in each age class and its standard deviation.

We estimated the body condition of adult female feral horses at the end of winter in September 2023 and 2024. The evaluation was made visually using the body condition score (BCS) of Rudman and Keiper (1991), ranging from zero to five, where zero is very thin and five is very fat. Following Bostal et al. (2024), we approached as many harem-bands as possible and randomly selected one or two adult females from each group, visually estimating their BCS while standing less than 30 m behind them.
Determination of survival and population growth rateFor the feral horse population of ETPP, the biological year runs from 1 November of year t, the median birth date, to 31 October of the year t + 1. A horse was assumed to have died if it was not registered in subsequent surveys within one year since its last sighting. For 1995–2002, we used mean survival rates and their standard deviations reported by Scorolli and Lopez Cazorla (2010). To calculate the survival rate for 2023 and 2024, we divided the total number of females at each age class in the November census of year t + 1 by the number recorded in the corresponding census of year t. The finite pre-breeding population growth rate (λ) was calculated by dividing the total number of females alive just before the birth pulse in year t + 1 by the total number of females alive at the same point in the year t (Caswell, 1989).
Carcass analysisTravel between harems during the population censuses was used to actively search for carcasses, following zigzag patterns to ensure thorough coverage of the area. The vegetation in the sampled area, consisting mostly of short grasses less than 20 cm tall, allowed very good visibility at distances of up to 20 m (Fig. 1). We recorded the geographic coordinates of each carcass found using a smartphone GPS during all the harem-band censuses described above. Animal deaths were classified as resulting from puma predation if bite marks (especially on the neck), large bone fractures, or skull fractures were observed following the criteria of Bank et al. (2002) and Turner et al. (1992). Covering of the carcass with plant remains, and their location in areas with tall grasses, shrubs, or in caves were taken as additional evidence. Approximate age of the horse at the time of death was assigned based on teeth eruption and replacement observed in photographs taken from the mandibles of each carcass (Martin, 2002). The approximate time since death in months was estimated based on the presence of flesh and analysis of the state of decomposition.
Statistical analysisSurvival rate for each age class between 1995–2002 and 2023–2024 was compared using a proportion test. We also assessed differences in the proportion of juveniles between 1995–1998, 1999–2002, and 2023–2024 using the same test. We used Chi-squared (χ²) tests to compare the proportion of adult females in poor (≤1.5), fair (2–2.5), or good (≥3) body condition (BCS) between 2023–2024, 2001–2002 and 2012–2013, the last two periods taken from Scorolli (2021). All statistical analyses were performed with a significance level of α = 0.05 in R (R-Core-Team, 2014).
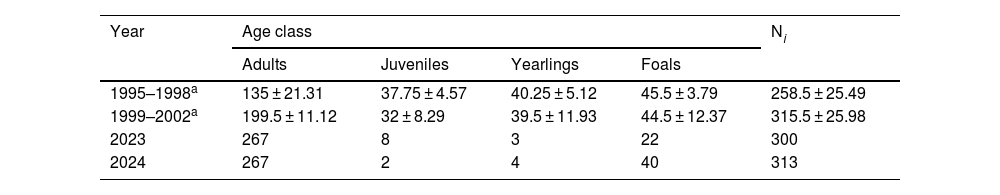
ResultsPopulation size of female feral horses was estimated at 300 in 2023 and 313 in 2024 (Table 1). Considering that the sex ratio of the feral horse population is usually 1:1, the population density for the current period can be estimated at 30 horses/km2.
Number of female horses by age class during 1995–2002 and 2023–2024 at Ernesto Tornquist Provincial Park, Argentina. Values for the period 1995-2002 are counts mean ± standard deviation and 2023-2024 annual counts.
| Year | Age class | Ni | |||
|---|---|---|---|---|---|
| Adults | Juveniles | Yearlings | Foals | ||
| 1995–1998a | 135 ± 21.31 | 37.75 ± 4.57 | 40.25 ± 5.12 | 45.5 ± 3.79 | 258.5 ± 25.49 |
| 1999–2002a | 199.5 ± 11.12 | 32 ± 8.29 | 39.5 ± 11.93 | 44.5 ± 12.37 | 315.5 ± 25.98 |
| 2023 | 267 | 8 | 3 | 22 | 300 |
| 2024 | 267 | 2 | 4 | 40 | 313 |
Ni: annual abundance of female horses.
Consistent with the change observed in the population structure, we noticed a decrease in foal and yearling survival in 2023–2024, coinciding with the presence of the puma. Foal survival rate was significantly lower (χ² = 154, p = 2.54 × 10−³⁵) in 2023–2024 compared to 1995–2002, while adults showed the inverse pattern (χ² = 21.2, p = 4.08 × 10−⁶). No differences were found when comparing this parameter for yearlings (χ² = 0.424, p = 0.515) and juveniles at two-years-old (χ² = 0.198, p = 0.657). Annual finite population growth rate (λ) was estimated at 0.975 and 0.982 for 2023 and 2024, respectively (Table 2).
Survival rate for different age classes of female feral horses during 1995-2002, 2023 and 2024 at Ernesto Tornquist Provincial Park, Argentina. Values for the period 1995-2002 are mean survival rate ± standard deviation.
| Year | Adults | Juveniles | Yearlings | Foals |
|---|---|---|---|---|
| 1995–2002a | 0.886 ± 0.026 | 0.907 ± 0.003 | 0.866 ± 0.052 | 0.936 ± 0.023 |
| 2023 | 0.978 | 0.750 | 0.667 | 0.182 |
| 2024 | 0.940 | 1 | 0.750 | 0.282 |
The proportion of juveniles in the population was 48% in 1995–1998, 37% in 1999–2002 and 12% in 2023–2024. The difference in proportion was highly significant between 2023–2024 and 1995–1998 (χ² = 81.4, p = 2.2 × 10−¹⁶), 2023–2024 and 1999–2002 (χ² = 50.2, p = 1.35 × 10−¹²), and significant between 1995–1998 and 1999–2000 (χ² = 5.1, p = 0.023).
The body condition of 119 adult females was assessed in 2023–2024 (Fig. 2). The proportion of mares at different body condition varied significantly between 2001–2002 and 2023–2024 (p < 0.01), while no significant differences were found when comparing 2012–2013 to 2023–2024 (p > 0.05).
, fair (2–2.5), and good (≥3) body condition scores, following the scale of Rudman and Keiper (1991), at the end of winter in 2001–2002, 2012–2013 and 2023-2024 at Ernesto Provincial Tornquist Park, Argentina. Data of 2001-2002 and 2012-2013 were taken from Scorolli (2021).")
Percentage of adult female feral horses with poor (≤1.5), fair (2–2.5), and good (≥3) body condition scores, following the scale of Rudman and Keiper (1991), at the end of winter in 2001–2002, 2012–2013 and 2023-2024 at Ernesto Provincial Tornquist Park, Argentina. Data of 2001-2002 and 2012-2013 were taken from Scorolli (2021).
We found 55 young horse carcasses between 2022 and 2024 (Table 3): 54% were foals at six months old or less, 35% were foals between six months and a year old, 9% were yearlings and 2% were juveniles aged two years or older. Most deaths (74%) were concentrated in spring and summer, when foals born were zero to eight months old, and 53% of the carcasses showed clear signs of puma predation (Fig. 3). In many of the remaining cases, the state of preservation of the carcasses was so bad that it was impossible to determine the cause of death. Only two of the twelve adult carcasses found showed signs of puma predation. The spatial distribution of carcasses coincided with the area most heavily used by feral horses at ETPP (Fig. 4).
Young feral horse carcasses detected at Ernesto Provincial Tornquist Park, Argentina.
| Year of death | Foals < six months | Foals ≥ six months | Yearlings | Juveniles | Total |
|---|---|---|---|---|---|
| 2022 | 2 | 1 | 2 (1) | 1 (1) | 6 (2) |
| 2023 | 8 (3) | 8 (3) | 1 (1) | 0 | 17 (7) |
| 2024 | 19 (13) | 11 (6) | 2 (1) | 0 | 32 (20) |
The number of carcasses that showed clear signs of predation by puma is indicated in parentheses.

 feral horse carcasses found at Ernesto Tornquist Provincial Park (ETPP), Argentina, from 2022 to 2024.")
Our study is the first to record and analyse the impact of puma predation on feral horse demography in South America. Although at a global scale there are reports of feral horses being preyed upon by pumas and wolves, most feral horse populations live under virtually predator-free conditions (Boyce and McLoughlin, 2021). In the United States, for example, while pumas coexist with horses in several regions, frequent and intense predation has only been documented in three sites in southwest deserts (Andreasen et al., 2021; Greger and Romney, 1999; Turner, 2015). In Argentina, pumas prey on wild camelids (Donadio et al., 2010) and other introduced herbivores like red deer, wild boar and sheep (Zanón Martínez et al., 2016), but it remains unknown if they have any effect on other feral horse populations in the country (Scorolli et al., 2019).
It is practically impossible to attribute the drastic drop in foal survival in 2023 to any cause other than predation by pumas. Despite intense surveys sustained since the nineties, we have no recent or historical evidence of infectious diseases affecting horses, frequent accidents (such as entanglement with fences), or generalized signs of undernutrition. Recent foal survival in our study area is within the range reported in the papers by Turner et al. (1992) and Greger and Romney (1999), and markedly below those observed in populations without predators (Grange et al., 2009). The apparent increase in the number of births in 2024 could be assigned in part to the little increase in the frequency of field sampling. In any case, and considering that some births and deaths may have remained undetected, the survival rate for younger individuals could very likely be lower than calculated, as discussed for the cases in the US (Greger and Romney, 1999; Turner et al., 1992). This new population scenario at ETPP coincides with the recovery of puma numbers in the region (Chimento and De Lucca, 2014). Although we could not assign a cause of death to all young and yearling carcasses we found, we suspect that the majority were likely the result of puma predation. There are no estimates about puma densities in our study area, but an increasing frequency of sightings has been reported and documented since 2023, including solitary individuals and a mother with cubs (ETPP personal communication). The effects of these puma numbers are perfectly consistent with those reported by Turner and Morrison (2001) for a feral horse population in western Nevada, where just between 3 and 9 pumas were responsible for low foal survival rates over 12 years. Based on these data about the survival of young horses, and considering that the proportion of adults in 2023-2024 at the study site is similar to those reported for other feral horse populations with predation (Greger and Romney, 1999; Turner et al., 1992), the change in the age structure of the population documented in our paper can also be fairly safely attributed to the effect of pumas.
Top-down limitation of wild equid populations has been reported for plains zebras (Equus burchelli) in Africa (Duncan et al., 2024; Grange et al., 2004; Owen-Smith and Mason, 2005), feral ponies in Spain (Lagos Abarzuza, 2013; Lagos and Bárcena, 2022) and Przewalski horses (Equus ferus przewalskii) in Mongolia (Van Duyne et al., 2009). Although the current study at ETPP shows a similar density to that of 2002, the estimated population growth rate is slightly lower (just below 1 for 2023–2024, compared to 1.06 for 1995–2002, Scorolli, 2021). Here we also observe consistency with the parameters estimated for other feral horse populations under intense puma predation (Greger and Romney, 1999), and a rate that is lower than those observed in populations without predation (Berger, 1986; Garrott et al., 1991; Linklater et al., 2004; Roelle et al., 2010). The λ value for 2023–2024, close to one, along with high foal mortality and direct evidence of puma predation, suggest a population limitation (sensuSinclair, 1989) by this large carnivore for the feral horse population in the study area. Although we do not know for sure when puma predation began to influence the local population dynamics, evidence of predation started in 2021, when foals stopped being detected shortly after being born (ETPP Park Rangers personal communication).
The fact that predation was concentrated during the foaling season supports the idea that pumas are selective predators, taking advantage of increases in the availability of vulnerable prey (Knopff et al., 2010). This preference for foals was reported for other feral horse populations under puma predation (Andreasen et al., 2021; Greger and Romney, 1999; Turner and Morrison, 2001). Contrary to Andreasen et al. (2021), who reported that cougars preyed on all age classes of a feral horse population in the western Great Basin and eastern Sierra Nevada, USA, at ETPP only a few carcasses of two-years-olds and adults showed signs of cougar predation. The low number of yearlings preyed by puma at our study site was probably a consequence of foal predation the previous year in this closed population. The carcasses were detected in lower grasslands (550 m asl), coinciding with the area of most intense use by wild horses in ETPP. The environment there is relatively homogeneous in its topography and vegetation, with no obvious signs of any pattern in the distribution of the carcasses. The area includes streams, some patches of tall grass, and clumps of invasive alien trees and shrubs, all of which favour the predatory activity of the puma (Donadio and Buskirk, 2016).
Predation of foals seems to have a positive impact on the body condition and survival of mares at ETPP. The lack of a significant difference in body condition score (BCS) between 2023–2024 (when horse density was 30 horse/km2) and 2012–2013 (when density was 17–19 horse/km2), compared to the difference observed between 2001–2002 (when density was 35 horse/km2) and 2023–2024, suggests that the loss of foals in recent years may have allowed females to allocate more energy to their bodies. A physiological trait selected during the domestication process of feral horses, unlike other ungulates, appears to be their ability to maintain stable reproductive rates even in situations of resource limitation, sacrificing mothers well-being and survival (Grange et al., 2009). The lactation period is the most energetically demanding phase of life for adult females of wild herbivores, during which lactating females expend 1.5 times more energy than non-lactating females (McNab, 2002; Parker et al., 2009). The interruption of lactation, the most energetically demanding phase of life for adult females of wild herbivores, may explain not only the improved mean BCS of adult females but also the significant difference in survival rates of this age class between 2001–2002 and 2023–2024. Changes in mean BCS may be alternatively explained by differences in primary production (Parker et al., 2009), but this seems to be a less plausible explanation for the increase in BCS observed in our study. Rainfall is the primary driver of grassland productivity at ETPP (Frangi et al., 1980); in 2001 and 2002, it was exceptionally high (1480 mm and 1244 mm, respectively). We can so assume that forage was more abundant in those years compared to 2023–2024 when rainfall reached average figures (ETPP, personal communication). Moreover, and despite similar population densities, the horse population in recent years has a greater proportion of adults, who consume more resources. All this reinforces the idea that it was the loss of foals, rather than environmental productivity, that influenced the body condition and survival of adult females. In the short term, greater survival of adults and an age structure biased towards this age class could increase herbivory pressure on grasslands. Although if the reported changes are sustained over time, they would most likely lead to a reduction in the total density of the species and so in an amelioration of its impact.
Direct management of large herbivores is commonly considered a strategy to mitigate the effects of their high densities, or in other words, their dominant biomass in ecosystems (Boyce and McLoughlin, 2021). However, this approach is often methodologically challenging and demands a significant investment. In Argentina, the government plays a prominent role in managing protected natural areas. The instability of state policies and the economy challenges sustained management plans that require long-term commitment and substantial resources. In turn, the management of a charismatic species, like horses, often elicits a strong negative response from some sectors of society (Scasta et al., 2020; Scorolli, 2018). In the ETPP, the only management of the feral horse population occurred in 2007–2008, when individuals were captured using mobile corrals and subsequently relocated to land owned by the navy, army, and a federal security force. However, this action faced criticism from horse protection advocates on social media, resulting in political reluctance to implement further management measures (Scorolli, 2018). Predator-prey interactions, where they occur, could be considered an ally to management strategies (Andreasen et al., 2021). In the case of ETPP, the feral horse population is currently at a high density and its effects are likely to be similar or worse after years of overpopulation (de Villalobos et al., 2011; de Villalobos and Zalba, 2010; Loydi et al., 2012; Zalba and Cozzani, 2004). In the long term, Puma predation may help reduce feral horse populations to levels consistent with the area's conservation objectives. In addition, incorporating this natural predator into future management plans for feral horse populations could reduce the number of horses that need to be captured and the frequency of interventions required to control their numbers.
FundingThis study was financed with grants from Consejo Nacional de Investigaciones Científicas y Técnica (CONICET) (Interannual Research Project PIP KA2 11220210100637CO) and Universidad Nacional del Sur (Research Group Project, PGI 24/B335).
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We are grateful to the authorities of the Ministerio de Ambiente de la Provincia de Buenos Aires for permitting the research and providing the facilities and to the rangers of Ernesto Tornquist Provincial Park for their help in many aspects of logistical support. We thank Dr. Sarah King (Colorado State University) for kindly proofreading the manuscript, and Dr. Alyson M. Andreasen (University of Nevada) for assisting with the confirmation of predator attacks in some of our cases and for sharing photographs from her archives.

