





Understanding the main factors determining the maintenance of native species in agroecosystems is of great importance to promote biodiversity-friendly practices. Here, we assessed the individual and interactive effect of local (management intensity, tree basal area, and tree diversity) and landscape (forest cover) predictors on the diversity of bees and social wasps in shaded cocoa agroforests located in the Brazilian Atlantic Forest. Using Malaise and attractant traps to survey these insects on 29 agroforests, we revealed that bee species number was positively associated only with forest cover, whereas tree diversity influenced bee species composition. In contrast, wasp species number enhanced with increasing management intensity, but only in agroforests inserted in landscapes with low forest cover, while no variable influenced the composition of social wasp species. These results reinforce the greater tolerance and ecological plasticity of social wasps, which can benefit from intensified management in structurally complex systems such as shaded agroforests. Nevertheless, bees tend to be more sensitive and dependent on forest cover at the landscape scale. Therefore, restoration efforts to increase forest cover in more deforested landscapes are urgently needed to ensure the high conservation value of shaded cocoa agroforests, especially for bee assemblages.

The conversion of forests into agricultural areas is considered the main threat to global biodiversity. This process leads to the loss of native species and, consequently, threatens the provision of ecosystem services and the functioning of ecological processes (Ramankutty et al., 2018; Watling et al., 2020). Therefore, preventing forest loss and increasing forest cover in degraded landscapes are considered essential principles to enhance biodiversity conservation (Riva et al., 2024). Species conservation can also be achieved by improving matrix quality (Perfecto and Vandermeer, 2010; Arroyo-Rodríguez et al., 2020). For instance, agroecosystems that maintain native tree species, such as shaded agroforests, can exhibit a vegetation structure similar to that of native habitats, therefore prone to support greater biodiversity and consequently preserving a wide range of wildlife species (Costa et al., 2021; Niether et al., 2020; Ferreira et al., 2020). However, each agroforestry system can have different local environmental conditions (e.g., vegetation structure and management intensity) and be embedded in different landscape scenarios (e.g., remaining forest cover), which can determine its conservation value (de la Mora et al., 2013; Ferreira et al., 2024a). Therefore, understanding the relative importance of local and landscape predictors for preserving species diversity in agroforestry systems becomes crucial to promote biodiversity-friendly agricultural practices in human-modified landscapes (Arroyo-Rodríguez et al., 2020).
In tropical countries, cocoa plantations are important agroforestry systems in terms of socio-economic-environmental benefits. This is the case of traditional cocoa (Theobroma cacao) plantations (hereafter, shaded cocoa agroforest) in southern Bahia, Brazil, where cocoa is grown under the shade of emergent trees, including species native to the threatened Atlantic Forest (Myers et al., 2000). Such a combination of native and cocoa trees increases the heterogeneity of vegetation structure, and can provide supplementary habitat for native forest-dweller species (Cassano et al., 2009). In fact, these agroforests are recognized for their high conservation value, as they are used as temporary or permanent habitats by different terrestrial and flying mammals (Faria and Baumgarten, 2007; Ferreira et al., 2020, 2025), reptiles, and amphibians (Cervantes-López et al., 2025).
Shaded cocoa agroforests can also be highly valuable for insect conservation, particularly for bees and wasps (Bos et al., 2007; Ferreira et al., 2024a; Nascimento et al., 2025). Although bees are not considered the main pollinators of this crop (Toledo-Hernández et al., 2017), both bees and social wasps can use these agroforests as supplementary habitats (Ferreira et al., 2024a). However, as cocoa plantations exhibit different vegetation structure mostly driven by contrasted management practices, the local condition can differ among agroecosystems, with subsequent influence on species persistence. For example, the reduction of shade tree richness in Indonesian cocoa agroforests (i.e., greater management intensification) resulted in local changes in terms of temperature, humidity, canopy openness and herbaceous extract, which subsequently lead to the reduction in bees and wasp’s diversity (Bos et al., 2007). As the availability of nesting substrate is important for determining the occurrence of bees and wasps (Araújo et al., 2021; Morato and Martins, 2006), especially for species that nest above ground, the structural vegetation complexity can also predict the diversity of these insects in agroforests. Finally, landscape forest loss can also limit the availability of resources, as food and nesting substrate for bees and social wasps (Ferreira et al., 2024a), and therefore, the effect of management intensity may depend on the remaining forest cover (i.e., an interacting effect). However, to our knowledge, no study to date has assessed how local characteristics associated with landscape composition determine the diversity of bees and wasps in these agroecosystems.
Here, we investigated the effect of local (i.e., management intensity, tree basal area, and tree diversity) and landscape (i.e., forest cover) variables on the number and composition of bee and social wasp species across 29 shaded cocoa agroforests from the northeastern Brazilian Atlantic Forest. We predicted that farms under greater management intensity would retain lower species number, since such practices will imperil the persistence of more sensitive species. In addition, shaded cocoa agroforests with greater tree diversity, higher basal area of shade trees and inserted in landscapes with higher forest cover possibly offer a greater diversity of floral resources, availability of nesting sites, and suitable habitat, respectively. As a result, we expect these characteristics to be positively associated with the number of bee and wasp species in these agroforests (Basset et al., 2012; Flores et al., 2018; Medeiros et al., 2019). We also predicted that agroforests with high management intensity, lower tree diversity, lower basal area of shading trees, and inserted in landscapes with lower forest cover would show high species composition dissimilarity, due to the loss of bee and wasp forest-dweller species and the dominance of generalist species commonly found in degraded environments (Ferreira et al., 2015; Souza et al., 2010). Since landscape context can modulate the effects of local management (Aycart-Lazo et al., 2025), we expect that the response of bee and social wasp assemblages to local characteristics will be weaker in landscapes with greater forest cover.
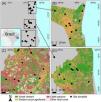
MethodsStudy areaWe conducted the study in the southern Bahia state, Brazil (Fig. 1) – where the economy has been based on cocoa production through shade agroecosystems (Faria et al., 2021), and other land cover types comprise native forests (i.e., Atlantic Forest), cattle pastures, eucalyptus monocultures, and urban areas (Mapbiomas, 2022). We selected 29 shaded cocoa agroforests distributed in three (Fig. 1) regions exhibiting different levels of deforestation and main land use types (i.e., low-deforested region = 54.1% of remaining forest cover, intermediate-deforested = 43%, high-deforested = 26.7%). The region with high-deforestation (∼4203 km²) is mainly dominated by cattle pastures (36.3%), following by forest remnants (26.7%), shaded cocoa agroforest (15.4%) and eucalyptus monoculture (4.6%); while the region with intermediate-deforestation (∼2181 km²) is dominated by forest remnants (43%) and shaded cocoa agroforest (33%). Finally, the region with low-deforestation (∼2,301 km²) is dominated by forest remnants (54.1%) and includes the most extensive remnant of Atlantic Forest (41,000 ha) in southern Bahia, under two protected areas, Una Biological Reserve and Una Wildlife Refuge.
Insect sampling
The sampling protocol is detailed elsewhere (Ferreira et al., 2024a), but a brief overview is given here. Within each agroforest, we established a 50-m transect located 100 m from the agroforest edge. Along this transect, we deployed three Malaise traps (Townes model) spaced 25-m apart, interspersed with six bait traps placed at ∼10-m intervals. The bait traps consisted of two with a water and sardine mixture, two with artificial orange juice, and two with artificial guava juice (protocol adapted from Souza et al., 2015). Sampling was carried out in a single campaign, for December 2022 to February 2023, during which all traps remained active continuously for approximately 72 h in each agroforest. The material was collected under license issued by the responsible Brazilian agency (ICMBIO license n° 83493-1) and specimens were deposited in the Invertebrates Collection of the Instituto Nacional de Pesquisas da Amazônia (INPA).
Local and landscape predictorsTo quantify the vegetation structure variables, we established four 50×50-m plots in each agroforest, separated by a minimum distance of 30 m. In these plots, we quantified, identified and measured the diameter at breast height (DBH) of all trees ≥ 10 cm (native and non-native species, including palm trees). Tree identifications were made at the lowest possible taxonomic level, with the assistance of an experienced botanist. Plant individuals not identified in the field were collected to be identified in the Herbarium of the Cocoa Research Centre at the Executive Commission of the Cocoa Farming Plan (CEPEC/CEPLAC), and the Herbarium of the Universidade Estadual de Santa Cruz (UESC). We then quantified two variables: the effective number of common tree species (i.e., Hill number in order 1, q1) using the iNEXT package (Jost, 2006; Hsieh et al., 2016), and the total basal area of native trees (m²/ha).
We performed interviews with farmers to quantify management intensity in each agroforest, obtaining information on four management practices: (i) frequency of weed control (per year); (ii) frequency of fertilization (organic or chemical) and/or liming (per year); (iii) frequency of pruning cocoa trees (per year), in which they often remove excess shoots and rarely carry out heavy pruning of larger branches and stems; in addition to directly counted (iv) the total number of cocoa trees in the established vegetation plot. The observed values were normalized by dividing each value by the highest observed value (separately for each variable) among all agroforests. The resulting values of the four variables for each agroforest were summed so that values equal to zero and four represent minimum and maximum management intensities, respectively (adapted from Mas and Dietsch, 2003).
Finally, we calculated landscape forest cover in multiple buffers (500, 750 and 1000 m) from each sampling site. To do this, we used a combination of two mappings: Mapbiomas collection 7 (Mapbiomas, 2022), which contains land cover but does not separate forest remnants from shaded cocoa agroforest, and Mapbiomas cocoa (Mapbiomas Cacau, 2023), which contains land cover from shaded cocoa agroforest but does not separate forest remnants from silviculture. Therefore, we used the combination of both mappings to only obtain the native forest cover in each landscape (i.e., forest cover). To determine the scale of forest cover effect (Jackson and Fahrig, 2015), we constructed Generalized Linear Models relating the response variable with forest cover measured at each scale. We then used the Akaike information criterion corrected for small samples (AICc; Burnham and Anderson, 2002) and identified that, for all response variables, the 500 m radii presented the best performance (ΔAICc = 0). For more details, see Appendices A, B and C in Supplementary Material 1.
Data analysisConsidering that bees and wasps can respond differently to habitat modification (Ferreira et al., 2024a), we conducted the analyses separately for each taxon. We used the iNEXT function from the iNEXT package to calculate sample coverage, and detected high variation among the sampled agroforests (range = 33 to 100%; Appendix B in Supplementary Material). For this reason, and given that our sampling effort was equal among all agroforests, we used species density per sampling effort (which corresponds to the observed number of species in each agroforest, hereinafter “species number”) as the response variable instead of estimated species richness. Changes in species composition across cocoa agroforests were quantified with the first axis of a Principal Coordinate Analysis (PCoA), calculated from a dissimilarity matrix using the vegdist function from the vegan package (Oksanen et al., 2022), constructed with a presence-absence matrix and Jaccard's index. To generate the dissimilarity matrix, we excluded one site where no individuals were collected, resulting in 28 agroforests. The first axis of the PCoA captured 27% of the variation in bee species composition, and 16% of social wasps. We build Generalized Linear Mixed Model and we use the model selection approach to assess the individual and interactive effect of local and landscape predictors on the response variables (number and composition species), and we include the “Region” (low, intermediate and high-deforestation) as a random factor. We did not detect multicollinearity among our explanatory variables (VIF < 3). For each response variable, we constructed eight models, which included each local variable individually (four models), the interactive effect of a local variable with landscape forest cover (three models), and the null effect (one model). We used a Poisson distribution for count data (species number) or a Gaussian distribution for continuous values (first axis of the PCoA). All fixed effect variables were standardized (subtracted by the mean and divided by the standard deviation) to zero mean and unit variance, ensuring that the coefficients represent changes in response per unit standard deviation of the predictor variable. Model selection was performed based on Akaike's criterion corrected for small samples (Anderson, 2008). We consider all models with ΔAICc < 2 to be parsimonious and, when more than one model was considered parsimonious, we selected the model with the highest Akaike weight. However, we selected the null model whenever it was present among the parsimonious models. We used the simulateResiduals function of the DHARMa package (Hartig, 2022) to assess whether that the residuals of our selected models did not violate the assumptions of homoscedasticity, dispersion and outliers (Appendix D in Supplementary Material). Subsequently, we performed a test for residual spatial autocorrelation (Moran’s I) using the testSpatialAutocorrelation function, from the DHARMa package (Hartig, 2022). We consider all results with a p-value < 0.05 to be significant. We did not detect any problems with model fit or residual autocorrelation. We conducted all analyses in R software (R Core Team, 2024).
ResultsWe collected 254 bees belonging to 38 species, and 346 wasps from 24 species (Supplementary Material 2). The average number of species of bees and social wasps per site was 4.3 ± 2.3 (range = 0–9 species) and 3.4 ± 2.1 (range = 0–10 species), respectively. Our model selection approach revealed that the number of bee species was best predicted by landscape forest cover alone (marginal pseudo-R² = 0.37; β = 0.25; p = 0.01), with a positive influence (Fig. 2A), whereas for the composition of bee species, the best model included only the shade tree diversity (marginal pseudo-R² = 0.20; β = −0.10; p = 0.03; Table 1 and Fig. 2B). Regarding the social wasp species number, the best model included the interactive effect of management intensity and landscape forest cover (marginal pseudo-R² = 0.29; β = −0.47; p = 0.01; Table 1). We observed that increased management intensity positively affected the number of social wasp species, but only in landscapes with low forest cover (Fig. 2C). Finally, for the composition of social wasp species, our model selection included the null model among the parsimonious models, and therefore we considered that none of the variables was able to explain the change in species composition (Table 1). The values for all models are presented in Appendix E in Supplementary Material.

Effect of predictor variables on the number of species of bees and social wasps in shaded cocoa agroforests in southern Bahia, in the Brazilian Atlantic Forest. We only show relationships from the selected models, shown in Table 1. In panel C, low (19%), medium (37%), and high (55%) values correspond to the first, second, and third quartiles, respectively, of forest cover in the landscape.
Parsimonious models (ΔAICc < 2) used to explain the species number and composition of bees and social wasps in shaded cocoa agroforests in southern Bahia, Brazil. When more than one model was considered parsimonious, we selected the model with the highest Akaike weight (Wi). The null model was selected whenever it was among the parsimonious models. The selected models are highlighted in bold.
| Taxon | Response variable | Model | k | ΔAICc | Wi |
|---|---|---|---|---|---|
| Bee | Species number | Forest cover | 3 | 0.00 | 0.60 |
| Species composition | Tree diversity | 4 | 0.00 | 0.42 | |
| Management index | 6 | 1.80 | 0.17 | ||
| Basal area * forest cover | 4 | 1.90 | 0.17 | ||
| Social wasp | Species number | Forest cover * management index | 5 | 0.00 | 0.71 |
| Species composition | Forest cover | 4 | 0.00 | 0.30 | |
| Management index * forest cover | 6 | 1.01 | 0.25 | ||
| Null | 3 | 1.07 | 0.12 |
In this study, we assessed the combined effects of local and landscape variables on bee and social wasp taxonomic diversity in shaded cocoa agroforests. As expected, we unveiled that bees and social wasps respond differently to habitat modifications at local or landscape scales. In particular, we demonstrated that only landscape forest cover explained the bee species number, but that the species composition was influenced by shade tree diversity. On other hand, only management intensity explained the wasp species number, but this effect was dependent on forest cover, while none of the variables explained the change in species composition. Considering that we are facing a global diversity and pollinator crisis (Betts et al., 2017; Levy, 2011), which calls for urgent management measures in human-modified landscapes (Arroyo-Rodríguez et al., 2020), our results demonstrate that both local management practices and landscape context should be considered to maximize bee and wasp species in cocoa agroforests.
Contrary to our expectations, none of the local variables explained the bee species number in cocoa agroforests. We attribute this low variation in the species number to the high heterogeneity characteristic of these agroforests, especially due to the presence of emergent and native Atlantic Forest trees (Cassano et al., 2009). In fact, in structurally more complex agricultural systems, bee species number may be similar to, or even higher than those found in native remnants within agricultural landscapes (Hoehn et al., 2010; Serralta-Batun et al., 2024), as also observed for these agroforests (Ferreira et al., 2024a). Furthermore, although the ages of the sampled agroforests unknown, this is a traditional system has expanded since the early 20th century and currently constitutes the main type of land use in this region (Faria et al., 2021). Therefore, considering the rapid life cycle of insects, it is possible that the bee assemblage present in these agroecosystems is composed of tolerant species to disturbance intermediate levels. Nonetheless, we observed that species composition was influenced by the shade trees diversity. This is to be expected, given that bees are highly dependent on floral resources (Michener, 2007) and that the occurrence of certain species may be determined by the availability of specific resources. Thus, greater tree diversity at the local scale may favor the occurrence of bees with more specialized relationships with host plants (González‐Chaves et al., 2024).
As expected, landscape composition proved to be important in determining the number of bee species in these agroforests. According to our predictions, landscape forest cover positively explained the increase in bee species number in surveyed agroforests, which can be explained by the habitat amount hypothesis (Fahrig, 2013). Landscapes with high forest cover may offer a greater availability and diversity of floral resources and nesting substrate, which can be accessed mainly by organisms that have a high vagility (Zurbuchen et al., 2010). Indeed, habitat loss has been one of the main causes of the decline of pollinators (Potts et al., 2010), including bees in agricultural landscapes (Saturni et al., 2016; Medeiros et al., 2019). Therefore, our results support the idea that maintaining or restoring forest remnants, especially in deforested landscapes, is an essential and priority measure to ensure the conservation of bee diversity in agricultural landscapes (Arroyo-Rodríguez et al., 2020; Riva et al., 2024), even in biodiversity-friendly systems.
For social wasps, we observed that the management intensity had a positive effect on species number, but only in landscapes with low forest cover. We attribute this response to factors such as the management practices adopted by cocoa producers and the resources provided by these agroforests, such as prey and nesting substrate. Our management intensity index includes practices such as pruning and thinning cocoa trees, which are less invasive than those employed in mechanized or intensive agricultural systems. These practices can also contribute to increase solar incidence, which may favor the activity of these wasps (da Silva et al., 2022) and increase prey density, typical of environments with more sunlight, such as forest edges and clearings (Barbosa et al., 2005; Pereira et al., 2017). Thus, as they are structurally complex agricultural systems, these agroforests may also provide an abundant supply of insects and nesting substrates (Ferreira et al., 2024b), on which social wasps are highly dependent (Barbosa et al., 2021). However, the positive effect of management intensity on the species number only in landscapes with low forest cover is unexpected, given that higher forest cover generally favors social wasp species richness in agricultural areas (Medeiros et al., 2019). In addition, it is important to note that the forest cover is negatively correlated with the amount of landscape agroforest (r = −0.77, p < 0.001). Therefore, less forest in the landscape implies an increase in the amount of supplementary habitat for these wasps, which may explain the positive response of these insects to intensified management in less forested landscapes. In this way, the environmental plasticity and generalist diet of social wasps, associated to the structural complexity of these agroforests, could favor these insects even under more intensive management.
The sustainability of agricultural practices depends on the adoption of strategies that minimize the trade-off between biodiversity conservation and agricultural yields. Despite the high conservation value of the shaded cocoa agroforests in the southern Bahia, this system has low productivity, considering the average production in other regions (Gama-Rodrigues et al., 2021). This lower economic gain can drive producers to adopt more intensive production methods with potential damage to biodiversity, like full-sun cocoa plantations or coffee monocultures, as previously observed in our region. However, studies suggest that the maintenance of shade trees associated with less invasive management practices (i.e., changes in the structural characteristics of agroforests such as the density of cocoa trees and associated trees), which is possibly the main bottleneck to greater productivity (Jagoret et al., 2017), can provide win-win scenarios for productivity and biodiversity conservation by enhancing the economic gains ensure the maintenance of high species number in these systems. In this context, our results indicate that agroforests inserted in landscapes with high forest cover can favor insect conservation, especially of bees, even under more intensive management. Additionally, these agroforests could experience greater productivity due to ecosystem services and ecological intensification associated, while maintaining local biodiversity (Araújo et al., 2025). Finally, although bees are not considered effective pollinators of cocoa, the pollination deficit is one of the factors associated with lower cocoa productivity (Toledo-Hernández et al., 2017). Thus, we can expect that maintaining a suitable habitat for bees (such as shaded cocoa agroforests), could also reflect in a favorable habitat for the maintenance of more effective cocoa pollinators (Toledo-Hernández et al., 2017). In addition, these benefits could also be extended to include the biological control offered by wasps and insectivorous vertebrates (Aycart-Lazo et al., 2025). Therefore, we suggest that restoration (in landscapes with low forest cover) or the maintenance of forest remnants (in landscapes with moderate or high forest cover), which could reduce the possible negative effects of intensified management on bee species richness, should be priority efforts to safeguard the taxonomic diversity of bees and social wasps in shaded cocoa agroforests.
Data availabilityThe datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We thank the farm owners for allowing us to conduct our research in their properties, as well as the team responsible for selecting the study areas (Martin Cervantes, Laís Rodrigues, Rebeca Sampaio, Paloma Resende, and Matheus Torres) and provided field assistance (Carla Rocha, Eduarda Caroline, Elimardo Bandeira, Júlia Ellen, Leonardo M. Abreu, and Maria Lavanholle). Finally, we would like to thank our colleagues from the Applied Ecology and Conservation Lab (LEAC) for their suggestions along the development of this study. This research was funded by The Rufford Foundation [nº 36668-1], the Brazilian Biodiversity Fund [FUNBIO, nº 039/2022], along with the Humanize Institute and Eurofins, umanize Institute and Eurofins, Idea Wild, and the Pró-Reitoria de Pesquisa e Pós-Graduação [PROPP nº 073.6764.2021.0013306-41 e nº 073.11010.2024.0029841-19] at the Universidade Estadual de Santa Cruz. José V. A. Ferreira and José A. dos Santos-Silva obtained a graduate scholarship [Finance Code 001] from the Brazilian Federal Agency for Support and Evaluation of Graduate Education (CAPES). Maíra Benchimol [304189/2022-7], Marcio L. Oliveira [311016/2023-5], José Carlos Morante-Filho [303302/2022-4] and Deborah Faria [303138/2022-0] obtained a productivity grant from the National Council for Scientific and Technological Development (CNPq). This study is part of the ‘Eco-nomia das cabrucas’ project (publication #20), coordinated by Deborah Faria and Maíra Benchimol, funded by the CNPq and the Research Support Foundation of the State of Bahia (FAPESB) [INC0006/2019], which support the National Institute of Science and Technology in Interdisciplinary and Transdisciplinary Studies in Ecology and Evolution (INCT IN-TREE).
The following are Supplementary data to this article:



