






Urban areas are expanding continuously inducing changes in raptor communities. However, our knowledge about urbanization impacts on raptor communities at continental scales is scarce. Therefore, the aim of this study is to relate geographical (city location) and environmental characteristics of cities with raptor species richness, taxonomic composition and functional composition in urban green spaces (UGS) of Neotropical cities. Data about raptor species present in UGS was compiled through search of scientific articles and thesis in databases. Raptor species richness increased in the tropics and decreased with increasing altitude. Species richness also increased with the increasing minimum UGS size of studies. Species composition dissimilarity was influenced by altitude and the geographic location of cities, and was dominated by species turnover. Scavengers were more frequent in cities of the east coast of Brazil, whereas semi-open habitat species were more frequent in southern South American cities. Raptor body size was related positively with UGS size. The results obtained confirmed the pattern of increasing raptor species richness in the tropics. However, anthropogenic factors such as green area loss in urban areas may influence the macrogeographical patterns of raptor species richness. The species composition dissimilarity between cities is dominated by a pattern of species replacements, related to species distributions in the Neotropics. Species functional traits were affected by macrogeographical factors and also by green area size, suggesting that urbanization may alter ecosystem processes provided by raptors. In short, the conservation and planning of large UGS is fundamental to maintain raptor communities in the Neotropical cities.
Urban areas are expanding continuously around the world, affecting several diversity facets of bird communities, such as taxonomic, functional and phylogenetic diversities (Ibáñez-Álamo et al., 2017; Morelli et al., 2017; Sol et al., 2017; La Sorte et al., 2018; Carvajal-Castro et al., 2021). Most of the studies were focused on Passeriform and Columbiform species, which are the dominant bird groups in urban areas (La Sorte et al., 2018). However, the analysis of raptor communities in urban areas is necessary due to their roles in ecosystem services such as carcass removal, regulating pest populations and cultural rituals (Şekercioğlu, 2006; Donázar et al., 2016; Kumar et al., 2019).
Several studies have shown negative impacts of urbanization on raptor taxonomic diversity and abundance (Bosakowski and Smith, 1997; Berry et al., 1998; Eduardo et al., 2007; Palomino and Carrascal, 2007; Sorace and Gustin, 2009; Hogg and Nilon, 2015, but see Rullman and Marzluff, 2014). However, urban green spaces (UGS) constitute hot-spots of bird diversity in urban areas due to their habitat heterogeneity, generally promoted by large UGS (Nielsen et al., 2014; Tryjanowski et al., 2017; Enedino et al., 2018, Amaya-Espinel et al., 2019; de Camargo Barbosa et al., 2020; Villaseñor et al., 2020). Large UGS have been found to increase raptor species richness in urban areas (Bosakowski and Smith, 1997; Hogg and Nilon, 2015). Nevertheless, it is unknown how cities and the characteristics of their UGS affect large-scale patterns of raptor communities in the Neotropics.
In the Neotropical region, raptor species richness has a latitudinal pattern, increasing in the tropics as a result of vegetation heterogeneity and low altitude (Diniz-Filho et al., 2002; Rangel and Diniz-Filho, 2003; Bellocq and Gómez-Insausti, 2005; Chapa-Vargas et al., 2019). Although the role of altitude has not been explored on raptors at large-scales in the Neotropics, other studies conducted on birds have shown species richness declines with increasing altitude (Rahbek, 1997; Herzog et al., 2005). On the other hand, the spatial patterns of raptor composition dissimilarity across the Neotropics have seldom been studied. Taxonomic dissimilarity patterns can be partitioned in turnover, which is the species replacement between areas, and nestedness, which is related to changes of species richness (Baselga, 2010). Studies conducted in several parts of the Neotropics have found that turnover patterns are the main form of spatial taxonomic dissimilarity of bird communities (Calderon-Patron et al., 2016; Fluck et al., 2020), which is related to geographic distance and changes in climatic conditions (Melo et al., 2009; Fluck et al., 2020). However, the roles of turnover and nestedness driving the spatial dissimilarity of raptor composition remain to be elucidated.
As top predators and scavengers, raptors provide several ecosystems services, regulating pest populations and removing livestock carcasses (Şekercioğlu, 2006; Donázar et al., 2016). A way to evaluate the anthropogenic effects on ecosystem services is through the analysis of functional traits (Dı́az and Cabido, 2001; Hooper et al., 2005), which indicate how resources are consumed by species (Dı́az and Cabido, 2001). However, the spatial composition of functional traits in raptors has been scarcely analyzed at large spatial scales (Huijbers et al., 2016; Sebastián-González et al., 2019). In the case of scavengers, climatic variables seem to influence their presence, although anthropogenic impacts seem to have a stronger impact on their presence (Huijbers et al., 2016; Sebastián-González et al., 2019). On the other hand, the effects of urbanization on raptor functional traits have been scarcely studied (Huijbers et al., 2015).
Therefore, the aim of this study is to relate geographical (city location) and environmental characteristics of cities with raptor communities in UGS. Specifically, it is expected that raptor species richness will increase in the tropics and decrease at high altitudes and, controlling for these effects, raptor species richness will increase with UGS size in cities. The taxonomic composition of raptors in cities will change mainly with geographical variables, showing a predominance of species turnover between cities, which will agree with natural species distributions in the Neotropics. On the other hand, raptor functional trait composition will change with geographical and environmental variables of cities. For example, scavenger raptors will increase their presence in the tropics, and diet and habitat specialist species will increase their presence with increasing UGS size.
MethodsLiterature searchScientific articles, theses and unpublished reports were searched during September 2015 (Leveau, 2021a), July 2020 and September 2021 in Google Scholar and Scopus (1960–2020), with the keywords [avian OR bird*] AND [green OR park OR cemeter* OR remnant* OR golf] AND urban, in English as well as in Spanish. Moreover, unpublished data obtained in urban parks of Buenos Aires city (Argentina) were included in the analysis (see Supplementary material, Table S1).
UGS were defined as patches dominated by vegetation and surrounded by a matrix of built-up land uses (Haq, 2011). This definition included urban parks, remnants of native vegetation, vacant lands, golf courses and cemeteries. Only papers that contained the following data were considered: (1) location of the study in the Neotropical Region (Morrone, 2014); (2) at least three UGS surveyed; (3) a list of the recorded species; (4) the temporal length of bird surveys; (5) the number of UGS surveyed; and (6) the minimum and maximum size of the UGS surveyed. Because some studies excluded raptors from their analysis, only studies that presented the record of at least one raptor species were included. Although studies conducted surveys for diurnal species, some nocturnal species also can be observed during the day. Therefore, this analysis included diurnal and nocturnal raptor species (Falconiformes, Cathartiformes and Strigiformes). Variables (4) and (5) were included in statistical models as covariates (see below) to control for sampling effort. The temporal length was classified as annual, where bird surveys were conducted during the breeding and nonbreeding seasons, and seasonal, where bird surveys were only conducted during the breeding or the nonbreeding season. Most of the studies conducted during one season were made during the breeding season. Annual surveys are expected to sample more bird species. On the other hand, the increased number of UGS is expected to increase the sampling of more raptor species. Other variables expected to affect the presence of raptors, such as latitude, longitude, altitude and city size were obtained from the sources or, when not available, from Wikipedia. Population size, a proxy of city area size (Ortman et al., 2014; Angel and Blei, 2016), was taken from the sources or, otherwise, from the national census conducted closest to the bird survey (5–10 years).
Functional traitsMain diet, habitat type and body mass (g) were obtained from bibliography (see Supplementary material, Table S2). Main diet and habitat type were obtained from the online source Birds of the World (http://birdsoftheworld.org). Diet was divided in: (1) generalist, that eat similar proportions of vertebrates and invertebrates and also carrion; (2) invertebrates; (3) birds; (4) mammals; (5) reptiles or fish; and (6) carrion. Habitat was divided in: (1) open, characterized by grasslands, deserts or aquatic environments; (2) semi-open, characterized by grassland with wooded areas; and (3) closed, characterized by a predominance of wooded habitats. Body mass was obtained from the EltonTraits database (Wilman et al., 2014).
Statistical analysisFor each study, the species richness and composition of raptors were obtained. Species richness was the number of species of each study. Species composition was the presence of the different species of raptors in each study.
The relationship between raptor species richness and the environmental variables (Table 1) was analyzed through generalized linear model (GLM). Due to the presence of overdispersion, a negative binomial model was performed with the function glm.nb of the MASS package in R (Ripley et al., 2019; The R Development Core Team, 2019). Multicollinearity was explored with Pearson correlations, and no strong correlations were found between environmental variables (r < 0.70). Models were obtained by backward elimination of non-significant variables (P > 0.05) from the full model using the anova function. Final models were compared with null models using a Likelihood Ratio test (LRT test) (P < 0.05). Plots of the regression models were constructed with the visreg package (Breheny and Burchett, 2017). Diagnostic plots of the final model were used to analyze homoscedasticity assumptions. The spatial autocorrelation of model residuals was analyzed with SAM software (Rangel et al., 2010). The residuals of the final model had not significant spatial autocorrelation (P > 0.05).
Continuous environmental variables used in this study. Also, the categorical variable Period was used (see “Methods”). Latitude and longitude are absolute values. masl: meters above sea level.
| Variable | Mean | Range |
|---|---|---|
| Latitude | 21.82 | 0.32−39.80 |
| Longitude | 63.25 | 34.83−98.73 |
| Altitude (masl) | 595.06 | 4−3625 |
| Number of green areas (N_GA) | 10.5 | 3−28 |
| UGS minimum area (Ha) (Min_area) | 5.14 | 0.01−50.00 |
| UGS maximum area (Ha) (Max_area) | 122.61 | 0.95−1000.00 |
| Population (inhabitants) | 1 811 630 | 29 433−8 800 000 |
The relationship between species composition and the environmental variables was analyzed through distance-based redundancy analysis (dbRDA). This method allow for carrying out constrained ordinations on data using non-Euclidean distance measures. In this case, a presence/absence based Bray-Curtis dissimilarity was calculated between studies. Species recorded in only one study were excluded. Then, the dissimilarity matrix and the environmental variables were related using the function capscale of the vegan package (Oksanen et al., 2017). Models were obtained by backward elimination of non-significant variables (P > 0.05) from the full model using the anova function. Final models were compared with null models using a Likelihood Ratio test (LRT test) (P < 0.05).
The partitioning of raptor composition dissimilarity between studies in turnover and nestedness was analyzed with the betapart package (Baselga et al., 2018). The Sorensen index of dissimilarity was partitioned with the function beta.multi, which compute the turnover and nestedness components for multiple sites.
The association between environmental variables and functional traits was analyzed through a fourth corner analysis (Brown et al., 2014). This analysis allows fitting a predictive model for species abundance (or presence/absence) as a function of environmental variables, species traits and their interaction. The function traitglm of the mvabund package was used (Wang et al., 2018). Because studies provided, in general, presence/abscense data of species, a binomial family was used to fit the model.
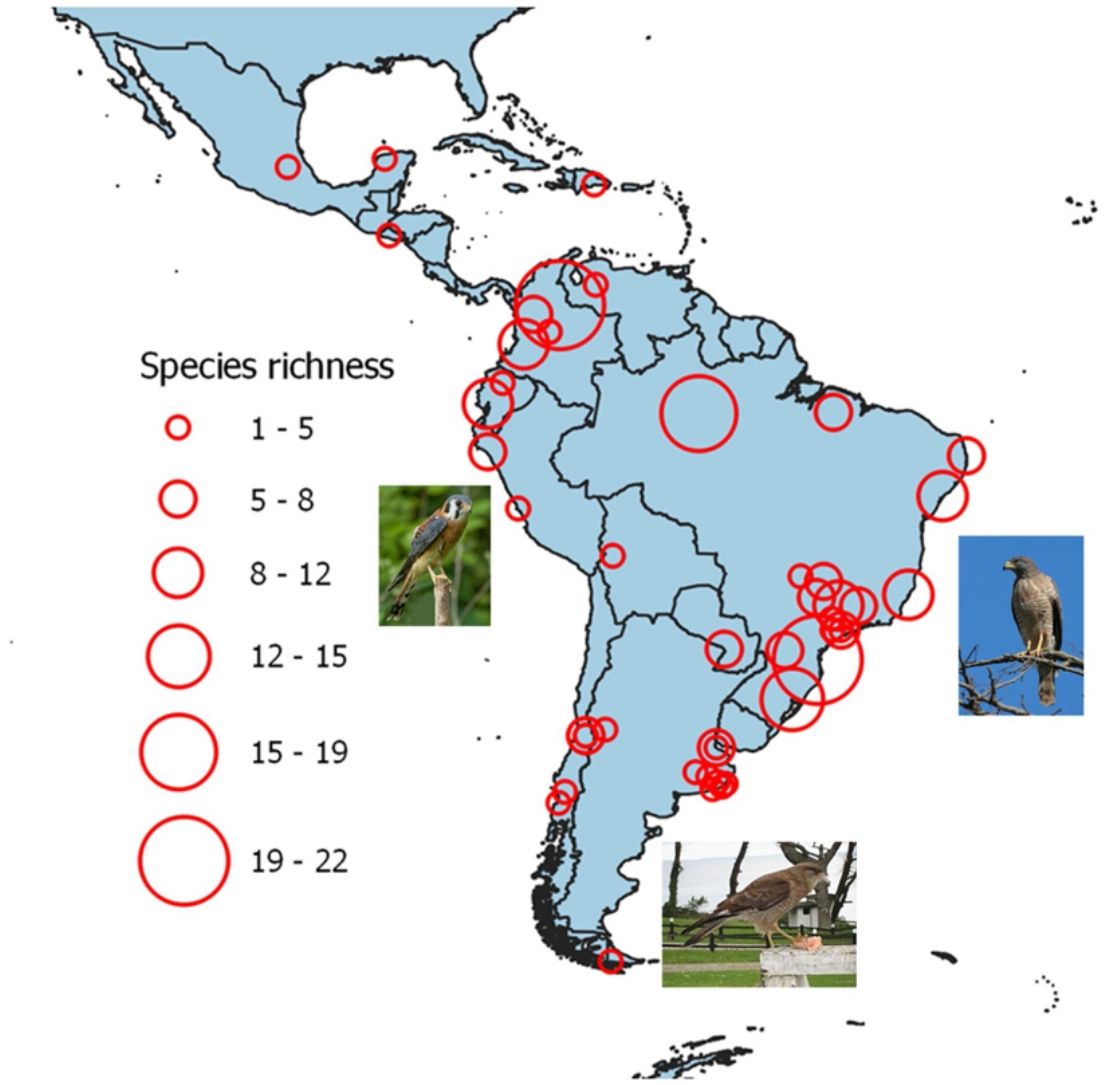
ResultsA total of 59 raptor species were observed in 49 studies belonging to 43 cities (Fig. 1), of which the Black Vulture (Coragyps atratus), the American Kestrel (Falco sparverius) and the Roadside Hawk (Rupornis magnirostris) were the most frequent species (Supplementary material, Table S3). The final model relating raptor species richness and predictor variables included latitude, altitude, minimum UGS size and period (LRT = 38.02, df = 4, P < 0.001; Table 2). Raptor species richness (mean = 5.53, range = 1–22) was negatively related to latitude and altitude, and was positively related to the minimum size of UGS (Table 2, Fig. 2). In addition, raptor species richness increased in those studies conducted annually in comparison to studies conducted during the breeding or nonbreeding season (Fig. 2).

Results of generalized linear model showing the relationship between the environmental variables and raptor species richness in urban green spaces (UGS) of cities in the Neotropical region. Period annual is in the intercept. masl: meters above sea level.
| Estimate | Std. Error | z value | P | |
|---|---|---|---|---|
| Intercept | 2.663 | 0.188 | 14.192 | <0.001 |
| Latitude | −0.040 | 0.007 | −5.697 | <0.001 |
| Altitude (masl) | <−0.001 | <0.001 | −2.616 | 0.009 |
| Minimum UGS size (Ha) | 0.024 | 0.007 | 3.557 | <0.001 |
| Period Seasonal | −0.510 | 0.189 | −2.700 | 0.007 |
-c), the red line represents the fitted curve and the grey areas are the confidence intervals at 95%. In d), the red line represents the mean values and the grey areas the confidence intervals at 95%.")
Generalized linear models showing the relationship between the environmental variables and raptor species richness in urban green spaces of cities in the Neotropical region. masl: meters above sea level. In a)-c), the red line represents the fitted curve and the grey areas are the confidence intervals at 95%. In d), the red line represents the mean values and the grey areas the confidence intervals at 95%.
The first two axes of the dbRDA explained 35% of the total inertia and showed that latitude, longitude and altitude were significantly related to raptor species composition in UGS (LRT = 7.70, P = 0.001). Longitude was related to the presence of the American Kestrel, the Variable Hawk (Geranoaetus polyosoma) and the Harris’s Hawk (Parabuteo unicinctus), and also related to cities located to the west part of Latin America, such as Santiago de Chile, Guayaquil, Quito and Lima (Fig. 3). Increasing latitude was related to the presence of the Caracara Chimango (Phalcoboenus chimango), and related to the southern cities located in central Argentina and Chile, such as Buenos Aires, Mar del Plata, Temuco and Valdivia (Fig. 4). Decreasing latitude and longitude was related to the presence of the Crested Caracara (Caracara plancus), the Roadside Hawk and the Black Vulture, and related to cities located in Brazil and Colombia, such as João Pessoa, Bogotá and Paragominas (Fig. 4). Increasing altitude was related to the presence of the American Kestrel, which was more frequent in cities near the Andes (Fig. 4).
 the relationship between the environmental variables and raptor species composition in urban green spaces in cities of the Neotropical Region, and (b) showing an expanded area delimited by discontinuous line in a). For species names, see Supplementary material, Table S3.")
Ordination analysis showing (a) the relationship between the environmental variables and raptor species composition in urban green spaces in cities of the Neotropical Region, and (b) showing an expanded area delimited by discontinuous line in a). For species names, see Supplementary material, Table S3.
 of (a) the Black Vulture (Coragyps atratus), (b) the Roadside Hawk (Rupornis magnirostris), (c) the Caracara Chimango (Phalcoboenus chimango) and (d) the American Kestrel (Falco sparverius) in cities of the Neotropics. Photo credits: Amestone1, Wagner Machado Carlos Lemes, hozdiamant and Greg Hume.")
Presence (filled red circles) of (a) the Black Vulture (Coragyps atratus), (b) the Roadside Hawk (Rupornis magnirostris), (c) the Caracara Chimango (Phalcoboenus chimango) and (d) the American Kestrel (Falco sparverius) in cities of the Neotropics. Photo credits: Amestone1, Wagner Machado Carlos Lemes, hozdiamant and Greg Hume.
On the other hand, the multiple site dissimilarity based on the Sorensen index was 0.96, of which 0.90 of dissimilarity between sites was due to species turnover and 0.06 of the dissimilarity was due to nestedness. Therefore, the latitudinal, longitudinal and altitudinal gradient of species composition is dominated by a pattern of species replacement between cities.
The fourth corner analysis showed associations between latitude, longitude, altitude, period of survey and size of UGS on raptor traits (Fig. 5). Scavenger diet increased toward the eastern tropics and in low altitudes, and also increased their presence with increasing minimum size of UGS. Species of semi-open habitats increased their presence in temperate areas. Species with a mammal diet increased their presence with increasing altitude and minimum size of UGS. Species of closed habitats were less frequent during surveys conducted along the year, whereas large raptors were more frequent in studies with the largest maximum area of UGS (Fig. 5).
 squares show positive relationships, whereas blue squares (−) show negative relationships. N_GA: Number of green areas; Min_size: minimum size of urban green spaces; Max_size: maximum size of urban green spaces.")
Fourth corner analysis showing the relationship between environmental variables and raptor functional traits in urban green spaces in cities of the Neotropical region. The scale bar indicates the interaction coefficients and the strength of the association between traits and predictor variables. Red values (+) squares show positive relationships, whereas blue squares (−) show negative relationships. N_GA: Number of green areas; Min_size: minimum size of urban green spaces; Max_size: maximum size of urban green spaces.
The results obtained confirmed the patterns found by other studies, with increasing raptor species richness in the tropics and decreasing species richness in high altitudes. Moreover, the results obtained also showed that green area size is important for raptor diversity in urban areas of the Neotropical Region, suggesting that anthropogenic factors such as green area loss in urban areas influence macrogeographical patterns of raptor species richness in cities. The taxonomic composition of raptors in cities was mainly related with large-scale variables, indicating that raptor species presence in cities is related with the regional pool of species surrounding each city. On the other hand, functional traits were associated with the geographical location of cities and also related to UGS size.
Patterns of species richnessThe results obtained indicated that raptor species richness increased in cities near the Equator and decreased with increasing altitude. A similar result was obtained through analysis of species map distributions (Rahbek, 1997; Diniz-Filho et al., 2002; Rangel and Diniz-Filho, 2003; Bellocq and Gómez-Insausti, 2005; Chapa-Vargas et al., 2019). Decreasing latitude and altitude are associated with more temperature and ambient energy, which promote more diversity of vegetation structure and, therefore, more raptor species richness (Bellocq and Gómez-Insausti, 2005; Chapa-Vargas et al., 2019). The increasing raptor richness in cities toward the tropics and in low altitudes suggests that the size of the regional pool of species influences the raptor community within cities, allowing the entering of more raptor species in the tropical cities.
Vegetation composition of UGS may change with latitude and influence raptor species richness. Several studies in the Neotropics suggest that tropical cities have more native trees than cities located in temperate areas, such as central Argentina, where tree species are exotic (Leveau and Leveau, 2006; Barbosa de Toledo et al., 2012; De Castro Pena et al., 2016). Therefore, more native vegetation species may favor the presence of raptors in UGS in the tropics.
The increasing minimum size of the UGS was positively related to the raptor species richness, suggesting that the UGS size also influence the presence of raptors in cities. The positive relationship between UGS size and bird species richness has been found to be strong and consistent worldwide (Leveau et al., 2019), and raptors in particular have shown positive relationships between species richness and the size of green areas in urban environments (Bosakowski and Smith, 1997; Hogg and Nilon, 2015). A decrease in green spaces area may be related to a reduced buffering against human disturbance, increased predation and a reduced prey density (Bosakowski and Smith, 1997; Schütz and Schulze, 2018).
More raptor species were detected in annual surveys than in surveys conducted during one season. This result agrees with the expectation that more survey effort would detect more species. Annual surveys can detect resident and also migrant species, such as the Osprey (Pandion haliatus) or the Broad-winged Hawk (Buteo platypterus) (Bierregaard et al., 2020a; Goodrich et al., 2020).
Surprisingly, city size did not affect the species richness of raptors. This could be related to the mobility of raptors, which can flight in large areas and probably are less affected by green spaces connectivity than other smaller birds, such as passerines. In fact, a recent analysis showed that migrant passerines species are affected by city size (Leveau, 2021a).
Patterns of taxonomic and functional compositionSpecies composition in UGS was mainly driven by the geographic location and altitude of cities. The dbRDA showed three main associations of species and cities in the Neotropics. Firstly, the Crested Caracara, the Roadside Hawk and the Yellow-headed Caracara (Milvago chimachima), whose distribution in the Neotropics is associated to lowlands of the tropics (Bierregaard and Marks, 2020; Bierregaard et al., 2020b, 2020c) and, in this study, their presence was mainly associated with cities of east Brazil, Colombia and Venezuela. Secondly, The Harris’s Hawk, the American Kestrel and the Variable Hawk, whose distribution in the Neotropics is associated to open and semi-open areas up to 5000 m (Dwyer and Bednarz, 2020; Smallwood and Bird, 2020; Bierregaard et al., 2020d) and, in this study, their presence was associated with cities near the Pacific coast and the Andes, such as Lima, Quito, La Paz and Santiago de Chile. Thirdly, the Chimango Caracara whose distribution is restricted to open and semi-open areas of the southern South America (Bierregaard et al., 2020e) and its presence was associated mainly to cities in the pampas of central Argentina and scrublands of Chile. These patterns of species distributions, where raptors species seem to replace one another across cities in the Neotropics, agrees with the partitioning of species dissimilarity between studies, where the species turnover component was dominant. Moreover, these results support the idea that raptor species presence in urban areas is related to their frequency in surrounding non-urban areas (Leveau et al., 2022).
The environmental variables analyzed, such as UGS size and population, were not significantly related to the taxonomic composition. This may suggest that raptor species are equally affected by green space size across the Neotropics, increasing their occurrence with increasing UGS size. On the other hand, population size of cities may not affect raptor species presence because their capacity for dispersal and movement. For example, raptor presence in big cities may be favored by the movement between several nearby green areas (Leveau, 2021b).
Functional composition of raptors was influenced by the geographic location of cities and the UGS size. Scavenger raptor presence in cities was negatively related to latitude and longitude, indicating more occurrences in tropical cities of Brazil, where the distributions of the Black Vulture, the Lesser Yellow-headed Vulture (Cathartes burrovianus) and the Turkey Vulture (Cathartes aura) overlap (Olrog, 1995). Moreover, scavenger species also were positively related with green space size, suggesting that the conservation of large green spaces may improve the presence of scavengers in urban areas. Raptors with a mammal diet were related to green spaces located in high altitude and with large minimum green space sizes. This result could be related to the distribution of the Variable Hawk in the Neotropics, whose presence range from lowlands to 3500 masl (Olrog, 1995). The main two mammal specialist species, the Variable Hawk and the White-tailed Kite, are medium to large sized raptors that may be area-sensitive species. Species of semi-open habitats increased their occurrence in southern cities. This pattern could be related to the presence of the Chimango Caracara in the Chilean and Argentinian cities located in southern South America. On the other hand, the presence of raptors with large body sizes increased with increasing maximum area of UGS. Large raptors generally have larger home ranges than smaller raptors, thus needing large UGS. Some large raptors, such as the Osprey and the Great Black Hawk (Buteogallus urubitinga), are associated to aquatic environments (Bierregaard et al., 2020a; van Dort, 2020), which generally are present in large UGS. Therefore, the results obtained suggest that raptor functional traits in the neotropical cities are influenced by macrogeographical and UGS characteristics.
ConclusionsCities seem to follow natural latitudinal gradients of raptor diversity, harboring more raptor species in the tropics. The size of UGS was an important driver of raptor diversity, highlighting the conservation and design of large UGS in cities. Raptor species composition was not related to characteristics of green areas or city size, but related to the location of cities in the Neotropics. Therefore, raptor species presence in cities may be more related to the regional raptor assemblages and the species that can colonize cities form the surrounding rural or natural areas. However, functional traits analysis showed that raptor body size was influenced by UGS size, suggesting that ecosystem processes provided by raptors are altered by the loss of green areas in cities. More studies are needed exploring the roles of UGS isolation and habitat vegetation characteristics driving raptor species richness and composition. On the other hand, the studies used in this analysis were not specifically designed to survey raptors, but instead entire bird communities. Therefore, future studies should design methodologies for raptor surveys to validate and expand the results obtained in this study.
Ethical approvalNot apply.
Funding sourcesThis research was funded by the Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación project number PICT-2015-0978.
CRediT authorship contribution statementNot apply.
Declaration of competing interestThe author declares that there is no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. Maria Alice Alves and two anonymous reviewers provided valuable comments that improved an earlier version of the manuscript.
I thank Latin American researchers who collected data used in this study.
The following are Supplementary data to this article:




