


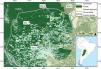



The distribution of the jaguar has decreased by approximately 50% with its conservation highly dependent upon its persistence and mobility in anthropogenic landscapes. Consequently, understanding the effects of land use on jaguar populations and their connectivity is a necessary precursor for effective conservation of the species. We simultaneously estimated jaguar density and landscape connectivity in the ranching landscape of the Dry Chaco of western Paraguay, a deforestation hotspot, as a function of proportional forest area using spatial capture-recapture modeling. Using camera trap sampling at four sites along a deforestation gradient of 17%–51% area deforested, we estimated densities of 0.44–1.6 individuals/100 km2, whereby densities and connectivity from the more deforested sites were significantly lower than those from the less deforested sites. Our results warrant concern for the long-term viability of jaguar in the Paraguayan Dry Chaco, highlighting the need for the effective implementation of existing national laws and management plans for the conservation of the jaguar and its habitat. Furthermore, we showed the importance of accounting for landscape heterogeneity typical of anthropogenic landscapes in the conservation of the jaguar, suggesting results from protected areas may be generating unrepresentative inferences for jaguars in general, while indicating the need to place a greater research emphasis on anthropogenic landscapes to meet range-wide conservation goals for the jaguar.
Habitat loss and fragmentation from expanding agricultural land use is a principle threat to biodiversity conservation (Foley, 2005; Haddad et al., 2015) as they negatively affect populations by reducing habitat availability, landscape connectivity, and the movement of individuals and genes (Fahrig, 2007, 2003). For mammals, extinction risk increases with increasing fragmentation and loss of connectivity, with increased risks for large terrestrial carnivores (Crooks, 2002; Crooks et al., 2017, 2011), which is evident in the high rates of extinction and decreases in distribution and abundance of these species worldwide (Ceballos, 2002; Ripple et al., 2014). Additionally, the negative effects on habitat availability and connectivity from agricultural expansion and the associated increased human footprint is exacerbated for large carnivores as they facilitate the hunting of carnivores and their prey, while also leading to increased persecution from conflicts with ranching interests or out of fear (Machovina et al., 2015; Ripple et al., 2014).
The implications of the loss of habitat and landscape connectivity from agricultural land conversion is evident for the jaguar (Panthera onca), the largest felid in the Americas and the apex predator throughout its distribution, whose range has contracted by ∼50% from its historic extent (de la Torre et al., 2018; Petracca et al., 2018; Thompson and Velilla, 2017). In response to the declining population and distribution of jaguars a range-wide conservation framework has developed that consists of the conservation of core populations (Jaguar Conservation Units; hereafter JCUs), largely dependent upon protected areas, linked by connectivity corridors within anthropogenic landscapes (Rabinowitz and Zeller, 2010; Sanderson et al., 2002; United Nations Development Program et al., 2019). Since most protected areas are insufficiently small to maintain viable populations in the absence of sufficient connectivity given the jaguars’ large spatial needs (Morato et al., 2016; Thompson et al., 2021), the survival and movement of jaguars through anthropogenic landscapes outside of protected areas is essential for the long-term survival of the jaguar (López-Bao et al., 2017).
Despite the contraction of the jaguar’s occurrence outside of protected areas (de la Torre et al., 2018), and anthropogenically driven loss of genetic diversity in jaguar populations throughout its range (Haag et al., 2010; Lorenzana et al., 2020, 2022; Roques et al., 2016; Srbek-Araujo et al., 2018; Wultsch et al., 2016), the large majority of research on jaguar population ecology has occurred within protected areas (Boron et al., 2016; Foster et al., 2020; Tobler and Powell, 2013). This lack of research focus upon anthropogenic landscapes in population estimation is of concern as the inferences generated from research in protected areas may not be representative of populations outside those areas and could lead to misinformed decision making (Allen et al., 2017; Tobler and Powell, 2013). Consequently, there is an inherent need for a shift in research on jaguar population ecology to focus more upon anthropogenic landscapes. This is especially true for areas outside of the Amazon where population declines have been greatest and where the conservation of remaining populations is highly dependent upon the jaguar’s movement through and survival in anthropogenic landscapes.
The negative effects of agricultural land use on the jaguar is most notable at the southern limit of its distribution where it has been extirpated from Uruguay, most of its range in Argentina, and the majority of the Atlantic forest of Brazil and Paraguay (de la Torre et al., 2018; Paviolo et al., 2016; Quiroga et al., 2014). Historically, the Dry Chaco of Argentina, Bolivia, and Paraguay has been important habitat for jaguar, and until recently the Dry Chaco of western Paraguay had been a population stronghold and connectivity hub for the species (Sanderson et al., 2002; Rabinowitz and Zeller, 2010; Thompson and Velilla, 2017). However, during the last twenty years, the Dry Chaco of Paraguay has become a deforestation hotspot, undergoing some of the world’s highest rates of deforestation, mostly driven by expanding livestock production.(Baumann et al., 2017; Caldas et al., 2015; Hansen et al., 2013; Vallejos et al., 2015), This deforestation has reduced the availability of preferred jaguar habitat, while increasing persecution and reducing prey availability, resulting in increased space use by jaguars and decreases in their abundance and population connectivity (Romero-Muñoz et al., 2020, 2019; Thompson et al., 2021, 2020; Thompson and Velilla, 2017).
As is typical of the majority of jaguar density estimates, population surveys of jaguar in the Dry Chaco have been conducted in relation to protected areas and indigenous reserves. In Bolivia, densities of 0.31–1.82 individuals/km2 were estimated (Noss et al., 2012), while in Argentina surveys showed that the jaguar is nearly extirpated in the region (Quiroga et al., 2014). Despite the extensive habitat loss and its apparent negative impacts on jaguars in the Dry Chaco (Romero-Muñoz et al., 2019), there have been no population surveys of jaguars in the region’s working landscapes, nor has there been any rigorously conducted surveys in the Paraguayan Dry Chaco or anywhere else in Paraguay. Moreover, there has not been an analysis of the effects of land use in the Dry Chaco on jaguar density and population connectivity.
Consequently, given the need to better understand the implications of widespread deforestation for the jaguar in general, and specifically in the Paraguayan Dry Chaco, we were interested in quantifying the effect of forest cover on jaguar density and population connectivity in deforestation hotspots, using the Paraguayan Dry Chaco as a case study. To do this we surveyed jaguars using camera trapping along a deforestation gradient in the Paraguayan Dry Chaco in a spatial capture-recapture (SCR) framework, taking advantage of modeling developments that allow for jointly estimating density and landscape connectivity (Morin et al., 2017; Royle et al., 2013; Sutherland et al., 2015). Hence, our objectives were to generate 1) the first spatially explicit density estimates for jaguars in Paraguay, and specifically for the Paraguayan Dry Chaco, and 2) the first estimate of the effects of habitat conversion on jaguar densities and population connectivity. Aside from important applications for the conservation of the jaguar in the Dry Chaco, our findings and the use of SCR modeling extensions have broader applications for understanding the effects of large-scale habitat loss and land management on jaguar density and population connectivity throughout the species’ distribution.
MethodsStudy areaThe Dry Chaco is the largest subtropical dry forest in the world, covering ∼787,000 km2 in Argentina, Bolivia, Brazil, and Paraguay (Olson et al., 2001), and is a global deforestation hotspot with 27% of its original forest cover transformed to mostly row crop agriculture and pastures (Caldas et al., 2015; Da Ponte et al., 2021; Hansen et al., 2013; Vallejos et al., 2015). About 22% of the Dry Chaco is in Paraguay, where 77,000 km2 (45%) of its forest area was lost by 2018, primarily driven by pasture creation for cattle raising, with an annual transformation rate of about 4% equating to a forest conversion rate of ∼500 ha/day/yr since 2000 (Baumann et al., 2017; Caldas et al., 2015; Da Ponte et al., 2021; Hansen et al., 2013; Vallejos et al., 2015).
Data collectionOur sampling was conducted in the Dry Chaco ecoregion of Paraguay in the departments of Alto Paraguay and Boquerón during 2016–2018, beginning during the austral fall or winter (May-September) and finishing during the austral spring, summer, or fall (September-May), dependent upon starting dates (Fig. 1, Table 1). We sampled four sites (Fig. 1) along a gradient of deforestation intensity, ranging from 17% to 51% deforested (see Data analysis), with trapping grids of 62–112 double camera trap stations (Table 1) located on roads, firebreaks, or along fence lines. All trapping grids had no public access and access to all study areas was provided by a single unpaved public road. No settlements were present throughout our study region and mean human population density per site was low, ranging from 0.1–0.5 people/km2 (Tatem, 2017). Within all study areas no hunting was permitted and we are confident that within the properties where we undertook sampling no jaguars were killed. However, at one site a GPS collared individual made a long-distance movement outside our study area where it was killed but we were able to control for this in our analysis. Although within our study sites no jaguars were killed, we confirmed illegal hunting at all sites (principally for gray brocket deer (Mazama gouazoubira) and collared peccary (Pecari tajacu)) despite prohibitions by ranch owners, however, based upon our observations and those of ranch employees it was not common.
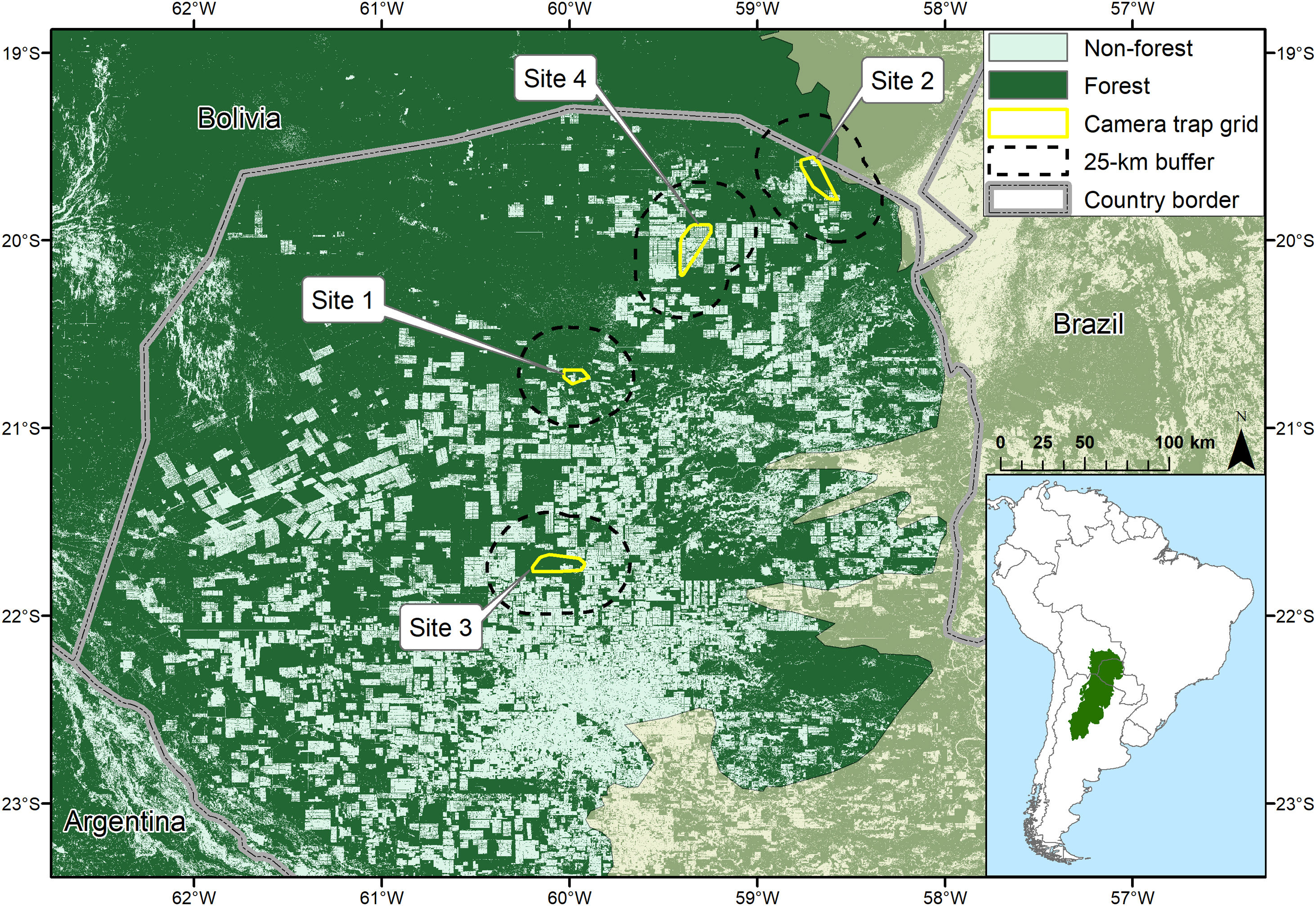
Location of the Dry Chaco in South America (inset) and locations of camera trap grids and their 25 km buffers in the Paraguayan Dry Chaco (Olson et al., 2001) in relation to forest cover in 2017 (Hansen et al., 2013).
Site-specific sampling characteristics of camera trap grids in the Paraguayan Dry Chaco, including sex-specific number of individuals captured and spatially recaptured (n), count of total captures, and count of spatial recaptures per site.
| Females | Males | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total captures | Spatial recaptures | Total captures | Spatial recaptures | |||||||||||
| % Deforested (25-km buffer) | State space area (km2) | Number of sampling stations | Sampling interval | Trap days | Mean operational days/station | n | count | n | count | n | count | n | count | |
| site 1 | 27 | 5503 | 86 | 18/08/2016–23/02/2017 | 9712 | 113 | 4 | 39 | 3 | 31 | 8 | 152 | 7 | 85 |
| site 2 | 17 | 8680 | 62 | 07/09/2016–18/01/2017 | 4038 | 65 | 4 | 38 | 4 | 17 | 9 | 47 | 4 | 29 |
| site 3 | 51 | 8470 | 112 | 27/05/2017–28/09/2017 | 12,552 | 112 | 4 | 100 | 4 | 47 | 2 | 97 | 2 | 50 |
| site 4 | 41 | 9085 | 75 | 30/08/2017–24/05/2018 | 8554 | 114 | 5 | 18 | 4 | 14 | 4 | 71 | 4 | 24 |
Due to the low detectability of jaguars and logistical considerations, sampling periods were long (124–279 days), with a mean of 65–114 operational days per station per site (Table 1). The site with the longest sampling period (Site 4) resulted from its sampling grid being divided into two temporally overlapping sections (48 and 27 stations). The long sampling periods were justifiable to increase detections (Dupont et al., 2019; Tobler and Powell, 2013), while we see no reason to expect any effect on density as a function of sampling year as there were no abnormal differences in temperature or precipitation between the sampling during 2016–2017 and 2017–2018 (Dirección de Metereología e Hidrología - Paraguay, 2022). Moreover, in our analysis we accounted for potential site-specific differences in detectability and space use (see Data analysis).
Minimum convex polygons of our trapping grids ranged between 80–282 km2; 0.7–2.5 times the size of mean telemetry-based estimates of 6-month activity areas (Krieged occurrence areas; Calabrese et al., 2016) of 12 jaguars (5 males, 7 females) in the Paraguayan Dry Chaco during the months corresponding to our sampling (McBride and Thompson, 2018; Morato et al., 2018a, 2018b; Thompson et al., unpublished data) and consequently were likely sufficiently large to minimize bias in density estimates (Sollmann et al., 2011; Tobler and Powell, 2013). Our modeling state space was defined by a 25-km buffer around our trapping grids, having a proportional area deforested ranging from 17%–51% (Hansen et al., 2013). As our study design was developed to estimate both community occupancy of medium and large mammals and jaguar density our camera spacing was lower than most jaguar-specific studies with mean distance between adjacent stations of 0.5–1.5 km.
Data analysisWe took advantage of extensions of the spatial capture recapture (SCR) modeling framework to explore the effect of forest cover on jaguar density and population connectivity in the Paraguayan Dry Chaco based upon our camera trap sampling data (Morin et al., 2017; Royle et al., 2013; Sutherland et al., 2015). By using the SCR ecological distance model the effect of landscape structure on animal space use is accounted for by employing a least-cost path distance (dlcp) predicated on a landscape covariate-specific resistance parameter (δ sensuSutherland et al., 2019; α2 sensuMorin et al., 2017; Royle et al., 2013; Sutherland et al., 2015). This allows for an estimation of how the landscape covariate affects individual space use as a deviance from the symmetric assumption of the traditional SCR Euclidian distance model (Morin et al., 2017; Royle et al., 2013; Sutherland et al., 2015). We considered this important as we expected that our highly structured landscapes and fragmented forest cover, along with the preferred use of forest by jaguars (Alvarenga et al., 2021; Morato et al., 2018a, 2018b; Thompson et al., 2021), would likely result in non-circular activity areas and spatial heterogeneity in detectability.
Using the resistance parameter (δ), the cost to movement across a landscape (dlcp), and the estimate of σ, the potential connectivity of a landscape (the expected frequency of use of a point in the landscape as a function of the cost to movement) can be estimated (Morin et al., 2017; Sutherland et al., 2015). Weighting the estimated potential connectivity by the estimated realized density (the number of activity centers estimated to be associated with a point in space) from the SCR model, allows for the estimation of the density-weighted (realized) connectivity whereby connectivity of a point in space in a landscape is quantified as a function of cost to movement and the distribution of individuals in the landscape (Morin et al., 2017; Sutherland et al., 2015). Both potential and density-weighted connectivity are comparable measures of landscape quality, allowing for the comparison among and within sites in space and time (Morin et al., 2017).
Using the photographic captures from our sampling we developed individual daily encounter histories for jaguars by assigning jaguar photographs to each sampling station by date and individual. Jaguars were identified by their spot patterns and sexed by genitalia, with juveniles accompanied by their mother excluded from the data set. Individual identification was undertaken independently by two researchers and any uncertainty in identifications clarified jointly by the researchers. We collapsed individual daily encounter histories to counts of the total number of daily detections per individual for each sampling period, assuming no temporal variation in detection during sampling periods, which facilitated computational efficiency. Using these spatial encounter histories we estimated densities for each site in a multi-site (multi-session) model in R (R Core Team, 2019) using the oSCR package (Sutherland et al., 2019) with a state space delimited by a 25-km buffer from traps at a resolution of 2 km.
We chose this resolution after an exploratory analysis of sensitivity in estimates to the resolution of the state space, finding that increasing resolution up to 2 km did not affect model estimates, while reducing computational time. Moreover, given that jaguars in the Paraguayan Dry Chaco can move in excess of 14 km per day on average (McBride and Thompson, 2018; Thompson et al., 2021) this resolution is consistent with the scale of jaguar space use in our study areas. We defined a state space of a 25-km buffer around each trap grid as it was the distance beyond which estimates for all parameters across all sites stabilized and because detectability at that distance for all sites approached zero, ensuring that individuals beyond that distance would not be included in our samples, while including the activity centers of all detected individuals.
As forest structure and canopy cover in the Dry Chaco are highly variable (Mereles, 2005), we chose to use the mean proportional area in forest as it serves as a biologically relevant metric that can be applied throughout the ecoregion to represent the availability of preferred habitat regardless of differences in forest structure and canopy cover. We developed maps of forest occurrence from 1 arc-second spatial data on forest canopy density corresponding to the year of our sampling (Hansen et al., 2013), classifying all areas with a forest canopy density of ≥15% as forest (based upon ground truthing and our familiarity with the study system), and then deriving the proportional forest area for each grid cell of our 2 km resolution state space. Our initial modeling of the effect of forest cover on the location of activity centers, however, resulted in issues with model convergence and unrealistic estimates of density and their variance, so we consequently confined our analysis to include the effects of forest cover on connectivity based upon ecological distance. We tested for differences in density, potential connectivity, density-weighted connectivity among sites using the difference between estimates at the 95% level while propagating estimation error adjusted for sample size (Lane et al., 2009).
We tested a set of 8 models that always included a site (session) effect on density, to meet closure assumptions, and on σ (scale parameter of decline in p with distance) to account for potential differences in space use among sites. As we were specifically interested in modeling connectivity and landscape structure on individual space use, the effect of proportional forest area on the resistance parameter δ was included in all models. Since we collapsed individual encounter histories into counts, we always included station-specific sampling effort as an effect on baseline detectability (p0), while testing the effect of sex on detectability as well. Additionally, for models including and not including the effect of sex on detectability, we also tested the additive effects of session and sex on σ. Model selection was based on Akaike Information Criteria (AIC) weights (Burnham and Anderson, 2002) and models with uninformative parameters (significant at <85% confidence level) excluded from the final model set (Arnold, 2010).
ResultsWe detected between 6–13 jaguars per site and one GPS collared individual from one site was censored in the analysis after it moved out of our sampling area and was killed. The ratio of males to females photographed ranged from 0.5 to 2.25 and the number of spatial recaptures per site was 38–116 (Table 1). Of the model set, three models contained 100% of the model weights (Table S1), with the model containing a site-specific effect on σ the highest ranked supported by 71% of model weights. The second and third highest ranked models included uninformative effects of sex on detection and σ (Table S1). The highest ranking model had a significant positive effect of effort on detection (βeffort = 0.013, SE = 0.001; Table S2) and a significant negative effect of proportional forest area on landscape resistance (βδ = −1.306, SE = 0.05; Table S2).
Based upon the highest ranking model, density estimates ranged from 0.44 to 1.6 individuals/100 km2, baseline detectability was estimated to be between 32% and 66%, and estimates of σ ranged from 1.0 to 1.9 km (Table 2). Detectability increased by about 0.3 percent per day of sampling effort and landscape resistance (δ) decreased by about 12% for each 10% increase in forest area, translating to fully deforested areas having a landscape resistance 3.7 times greater compared to fully forested areas. Sex ratio estimates had high uncertainty, but the estimated probability of being a male varied from 36% to 70% across sites (Table 2).
Estimates and 95% confidence intervals (CI) from the highest ranking model for density, baseline detectability (p0), scale parameter (σ) by site, sex ratio (expressed as the probability of an individual being a male), and standardized landscape resistance of modeled state spaces.
| Density (Individuals/100 km2) | Detectability (p0) | σ (km) | Sex ratio (Probability of being a male) | Standardized landscape resistance | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| estimate | 95% CI | estimate | 95% CI | estimate | 95% CI | estimate | 95% CI | mean | 95% CI | |
| Site 1 | 1.26 | 0.70–2.27 | 0.39 | 0.30–0.49 | 1.4 | 1.2–1.6 | 0.68 | 0.38–0.87 | 0.11 | 0.00–0.58 |
| Site 2 | 1.60 | 0.90–2.83 | 0.39 | 0.29–0.49 | 1.0 | 0.9–1.2 | 0.70 | 0.42–0.89 | 0.06 | 0.00–0.56 |
| Site 3 | 0.58 | 0.26–1.29 | 0.32 | 0.24–0.40 | 1.3 | 1.2–1.4 | 0.36 | 0.85–0.73 | 0.23 | 0.00–0.71 |
| Site 4 | 0.44 | 0.23–0.85 | 0.66 | 0.49–0.80 | 1.9 | 1.6–2.1 | 0.44 | 0.17–0.75 | 0.24 | 0.00–0.81 |
Site-specific estimated densities, potential connectivity, and density weighted connectivity showed a general negative relationship with increasing deforested area (Fig. 2). Estimated density between the two least deforested sites (sites 1 and 2) were not significantly different at the 95% level, nor was the difference between the most deforested sites (sites 3 and 4; Fig. 3a) significant. However, estimated density for the least deforested sites (sites 1 and 2) were each significantly different from each of the most deforested sites (sites 3 and 4; Fig. 3a). The mean values of estimated potential connectivity were significantly different among all sites with the exception being between the two most deforested sites (sites 3 and 4; Fig. 3b), while the differences in the mean density-weighted connectivity were significantly different among all sites (Fig. 3c).
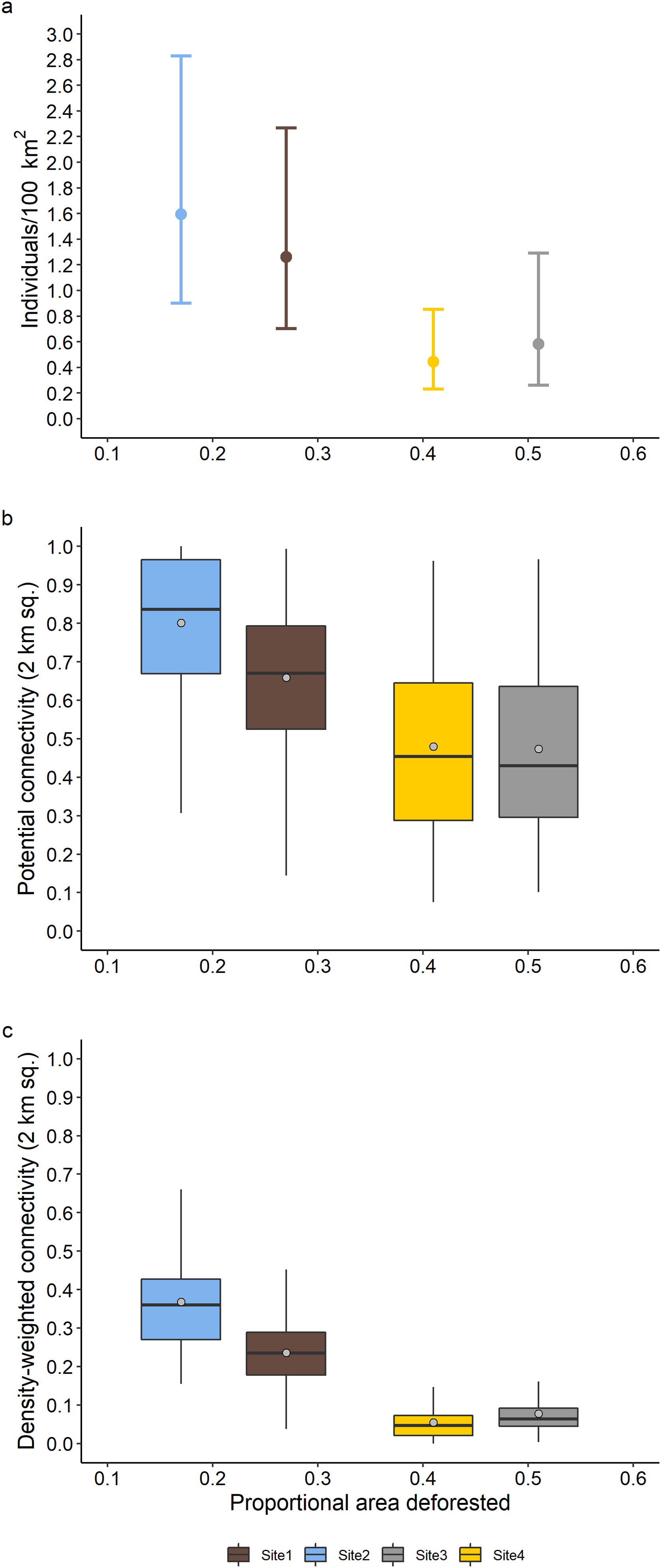
The relationship of proportional area deforested within the modeled state spaces with (a) density (error bars represent the 95% confidence interval), (b) standardized potential connectivity, and (c) standardized density-weighted connectivity. Boxes show the 25th to 75th percentile, vertical lines within the boxes the median, and the whiskers 1.5 times the interquartile range. Points in the boxes represent the mean value.
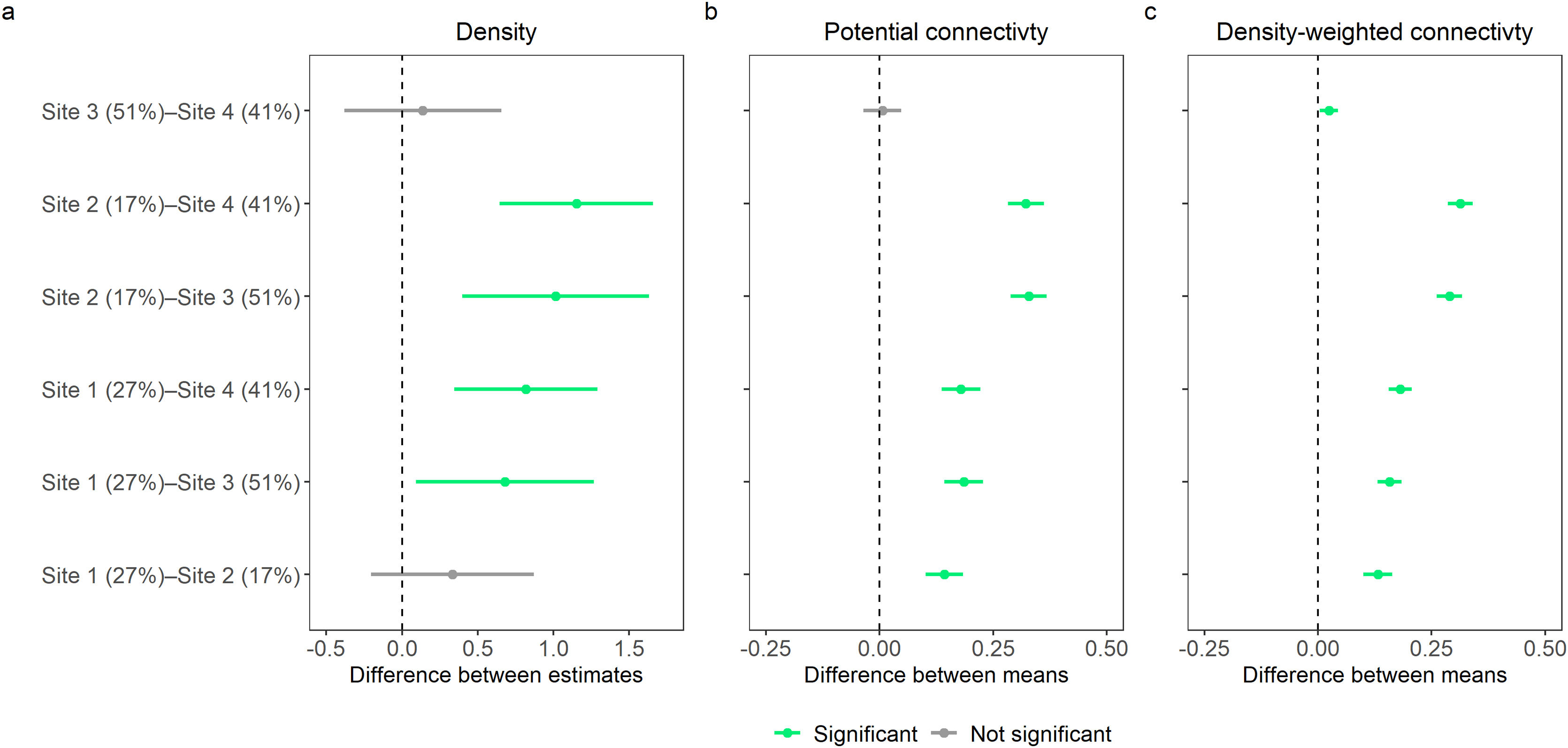
Difference between site-specific (a) density estimates, (b) mean estimated potential connectivity, and (c) mean estimated density-weighted connectivity. Points represent differences and bars represent the 95% confidence intervals. Where confidence intervals do not include 0 differences are significant. Percentages associated with site names represent the percent area deforested for state space of each trapping grid.
By taking advantage of newly developed modeling approaches and sampling across a gradient of deforestation intensity, our study is the first to examine the effect of landscape transformation on jaguar density and connectivity, apart from producing the first rigorous estimates of jaguar density for Paraguay. More generally, despite the recognized importance of connectivity for the long-term persistence of species (Lindenmayer et al., 2008), to date few studies have simultaneously estimated density and connectivity (Fuller et al., 2016; Pal et al., 2021), and this study is the first to do this across differing landscape structures. Despite the uncertainty in our density estimates we were able to show that estimated densities in the more deforested sites were significantly lower compared to densities in the less deforested sites. Moreover, by incorporating ecological distance in the SCR formulation, we found a strong dependence of connectivity and jaguar space use on forest area which was consistent with observed patterns in jaguar spatial ecology (Alvarenga et al., 2021; Morato et al., 2018a, 2018b; Thompson et al., 2021).
We did not find sex-specific differences in detectability or for σ which was contrary to other jaguar studies (Boron et al., 2016; Sollmann et al., 2011; Tobler et al., 2018, 2013) and not expected since space use and movements by male jaguars in the Dry Chaco, and range-wide, is considerably larger than that of females (McBride and Thompson, 2018; Morato et al., 2016; Thompson et al., 2021). The lack of sex-specific differences in detectability and σ could be explained by variations in individual space use and differences in sex ratios among sites offsetting the frequency of males and females detected across sites (Table 1). Estimates of σ were smaller than other studies (Boron et al., 2016; Sollmann et al., 2011; Tobler et al., 2013) which is attributable to our accounting for habitat-driven asymmetry in space use (Tobler et al., 2018). This was expected since the ecological distance formulation of the SCR model estimates smaller values of σ compared to the Euclidean distance formulation as the effect size of δ increases (Morin et al., 2017; Royle et al., 2013; Sutherland et al., 2015).
Jaguar occurrence in the Dry Chaco is subject to complex source-sink dynamics, driven by habitat loss, high anthropogenic mortality, and prey depletion (McBride and Thompson, 2018; Romero-Muñoz et al., 2020, 2019; Thompson et al., 2020). Consequently, our estimates of density and connectivity in relation to forest cover result from an interplay of the negative effects of high anthropogenic mortality and prey suppression acting synergistically with habitat loss. We found that connectivity in the more transformed landscapes is maintained, although greatly reduced, suggesting that populations can be augmented through immigration and that our study sites are potentially serving as populations sinks. Moreover, the rapid rate of forest loss in the Paraguayan Dry Chaco, and the longevity of jaguars, raises additional concerns that we underestimated the negative population effects of habitat loss as the true effect could be masked via a time lag (Tilman and Lehman, 1994) as has been observed for other mammals in the Dry Chaco (Semper-Pascual et al., 2018). The uncertainties over the degree of demographic influence of these mechanisms point to the need for long-term monitoring of jaguar populations to estimate vital rates and immigration, however, there are conspicuously few such estimates for jaguars (Glennie et al., 2019; Gutiérrez-González et al., 2015; Harmsen et al., 2017). Given high anthropogenic mortality and evidence of source-sink dynamics for the jaguar in the Dry Chaco (Romero-Muñoz et al., 2019), long-term monitoring of jaguars should be a priority for their conservation in the region, as well as for the species throughout its distribution.
In Paraguay the jaguar is listed as critically endangered due to its country-wide range constriction and habitat loss, with the majority of its remaining distribution being in the Dry Chaco (Giordano et al., 2017). Given the expected continuation of habitat loss, the relatively small amount of protected area in the region in relation to the jaguar’s spatial needs, and high levels of human-caused mortality (McBride and Thompson, 2018; Romero-Muñoz et al., 2019), the negative effects of deforestation on jaguar density and connectivity that we observed are of concern for the long-term conservation of the species in the Paraguayan Dry Chaco. This concern is further warranted as land use trajectories and jaguar population declines in the Dry Chaco are mirroring those that have led to the near extirpation of jaguars in the Atlantic Forest of eastern Paraguay, whereby extensive deforestation for agricultural production has confined the occurrence and movements of the remaining jaguars to protected areas and adjacent habitat patches with no connectivity among other populations (McBride and Thompson, 2019; Paviolo et al., 2016).
Within the Paraguayan Dry Chaco landowners are legally permitted to clear between 50% to 75% of forest area within their properties (Congreso de la Nación Paraguaya, 1973; Secretaria del Ambiente de Paraguay, 2001) and subsequently, our findings suggest that if private land is fully developed, the landscape outside of protected areas will be at best marginally suitable to maintain jaguars and likely serve as population sinks, while protected areas will be increasingly isolated (Romero-Muñoz et al., 2019). However, we showed that connectivity in our more transformed landscapes, albeit reduced by increasing forest loss, can be consciously maintained and we urge the enforcement of national forest conservation laws, the application of landscape planning, and the development of initiatives to incentivize forest conservation to achieve that. In addition, the high anthropogenic mortality of jaguars in our study area points to the need for habitat protection to be undertaken concurrently with efforts to reduce anthropogenic mortality. As jaguar killings are rooted in real or perceived livestock losses from depredation, enforcing and implementing relevant sections of the national jaguar conservation law (Congreso de la Nación Paraguaya, 2014) and management plan (Secretaría del Ambiente et al., 2016) to foster coexistence between the ranching community and jaguars would aid in reducing the negative effects from habitat loss through increasing jaguar survival on private ranchlands.
The anthropogenic context of jaguar conservation in the Paraguayan Dry Chaco is typical of jaguar populations throughout the species’ range and demonstrates the broad relevance of our findings as they show how jaguar density and connectivity relate to habitat loss in anthropogenic landscapes. Moreover, as jaguars exhibit strong spatial associations with forest cover and rivers (Alvarenga et al., 2021; Eriksson et al., 2022; Morato et al., 2018a, 2018b; Thompson et al., 2021), the strong asymmetrical habitat-driven space use that we found highlights the need to consider the potential negative bias induced in SCR modeling from assuming symmetrical space use when estimating jaguar density in heterogeneous landscapes, as well as the value of formally estimating connectivity within the SCR modeling framework (Morin et al., 2017; Sutherland et al., 2015; Tobler et al., 2018). Importantly, as the large majority of jaguar population studies have been in protected areas (Boron et al., 2016; Foster et al., 2020; Tobler and Powell, 2013), our results show that these studies are potentially generating skewed inferences for conservation as they fail to account for the complex spatial dynamics of populations in anthropogenic landscapes.
The case of the jaguar in the Paraguayan Dry Chaco illustrates the importance of understanding the drivers of jaguar population ecology in the landscape matrix outside of protected areas for the species’ long-term survival. Moreover, our findings demonstrate that adequately implementing range-wide initiatives for jaguar conservation (United Nations Development Program et al., 2019) will require a focus on quantifying how land use affects populations and movements of jaguars in anthropogenic landscapes, as well as on interventions promoting coexistence and habitat conservation, to achieve positive conservation outcomes in anthropogenic landscapes in Paraguay and across the jaguar’s range.
Author’s contributionsJ.J.T., M.V., J.M.C.K. and J.L.C. conceptualized the project and designed the methodology; J.J.T., M.V., V.R.B. administered the project; J.J.T., M.V., H.C., N.C., V.R.B., E.B., J.M.C.K., R.T.M., and R.A. collected the data; J.J.T. and N.C. curated data; J.J.T. analyzed the data; J.J.T., M.V., and J.L.C. led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Conflict of interestsThe authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We thank the ranch owners, administrators, and employees who facilitated and made our research possible. This research was funded by the Consejo Nacional de Ciencia y Tecnología (CONACYT) of Paraguay under project 14-INV-060. JJT, MV, HCB, VR, JMCK, and JLC were partially supported by CONACYT through the program PRONII. We thank Alberto Yanosky for supporting our research, Estebán Vasconcellos for his cooperation and logistical support, Ivan Benitez, Victor Robles, Javier Ovelar, Carlos Molinas, Patricia Salinas, Paola Cuevas, Johanna Lopez, Christian Palacios, Magdalena Benitez, Agustin Paviolo, José Aguilera and José Méndez for assistance with data collection, and Chris Sutherland for modeling advice.
The following is Supplementary data to this article:

Present address: Programa de Pós-Graduação em Biologia Animal, Universidade Estadual Paulista, 15054-000, São José do Rio Preto, SP, Brazil.
Instituto de Investigación Biológica del Paraguay.
Present address: Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida, USA.
Present address: Department of Large Animal Clinical Sciences, College of Veterinary Medicine, University of Florida, Gainesville, Florida, USA.


- Home
- All contents
- Publish your article
- About the journal
- Metrics
- Download PDF
- Bibliography
- Additional material