





Wildlife population surveys are crucial for conservation but require significant time and resources, especially for elusive species like the threatened maned sloth (Bradypus torquatus). We evaluated the efficiency and effectiveness of thermal drones versus traditional ground-based surveys for detecting and estimating maned sloth populations, across seven 10.5-hectare forest plots in Bahia, Brazil. Thermal drones detected 10 individuals in six plots (86%) over 13.8 h, while ground-based surveys detected nine individuals in three plots (43%) over 51.7 h. Detection rate were significantly higher for thermal drones (0.70 ± 0.45 individuals/hour) than ground-based method (0.17 ± 0.22 individuals/hour; p = 0.006), though density estimates were similar. Drone survey costs were initially four times higher but had lower operational costs per area (2.7×) and per sloth detected (3.2×), achieving financial return after surveying 740 ha or detecting 38 sloths. Drones were three times faster for individual detection and twice as efficient for identifying occupied sites, reducing costs and improving results. However, combining drones with ground surveys or increasing sampling effort with drones is recommended for accurate density estimates. This technology may be particularly advantageous for medium- to long-term studies and large-scale surveys, enhancing population monitoring and conservation of arboreal mammals like the maned sloth.

Arboreal mammals are highly vulnerable to forest loss and degradation, which impact their habitat and populations (Whitworth et al., 2019). Estimating population parameters such as occupancy, abundance, and density is crucial for understanding their responses to anthropogenic pressures and improving conservation strategies (Johnson, 2008; MacKenzie et al., 2002). However, detecting and counting arboreal species in tropical forests is challenging due to the difficulty in accessing the canopy (Haysom et al., 2021; Kays et al., 2019). Folivorous arboreal mammals, reliant on camouflage for predator avoidance, are particularly difficult to locate (Eisenberg, 1978; Giné et al., 2015), and many do not respond to playback or baits, limiting the effectiveness of traditional survey methods (Chiarello, 2008; Pocknee et al., 2021).
Unmanned Aerial Vehicles (UAVs, hereafter drones) equipped with thermal infrared (TIR) cameras offer a promising alternative for detecting and counting arboreal mammals in forest canopies (Kays et al., 2019; Spaan et al., 2019; Witt et al., 2020; Rahman et al., 2025; Ryan et al., 2025). Thermal drone surveys have outperformed traditional methods for detecting species like koalas and primates (de Melo, 2021; Howell et al., 2021; Spaan et al., 2019; Witt et al., 2020), often delivering higher detection efficiency, improved accuracy in population estimates, and greater cost-effectiveness over extended monitoring efforts (Howell et al., 2021; Spaan et al., 2019; Wearn et al., 2023; Witt et al., 2020). Thermal drones overcome detection challenges, such as animal camouflage and nocturnality, by identifying animals through body heat (Burke et al., 2019; Kays et al., 2019). Moreover, they can quickly scan large areas, facilitating repeated sampling and data collection even for species with low population density (Linchant et al., 2015; Ryan et al., 2025; Witt et al., 2020). However, their efficiency and accuracy vary with species, climate, technology, and vegetation density (Kays et al., 2019; Linchant et al., 2015; Rahman and Rahman, 2021; Ryan et al., 2025) and require validation through ground-truthing (Rahman et al., 2025).
Three-toed sloths (Bradypus spp.) illustrate these detection challenges particularly well. They exhibit extreme folivory-related adaptations, including an exceptionally low metabolism and body temperature (Cliffe et al., 2018), resulting in very slow movements that hinder detection by humans (Eisenberg, 1978; Lopes et al., 2023). They carry out their activities within the dense and tall canopy (Falconi et al., 2015; Montgomery and Sunquist, 1978), further limiting detection (Giné et al., 2015; Lopes et al., 2023). These cryptic habits hinder accurate population estimates and contribute to the scarcity of quantitative data on sloth populations (Chiarello, 2008). Traditional survey methods typically involve visually inspecting the canopy during the daytime along transects or within defined sampling areas (Garcés-Restrepo et al., 2019; Santos et al., 2019). This approach is labor-intensive and time-consuming (Santos et al., 2019, 2023), hindering large-scale investigations, repeated sampling, and accurate estimation.
The northern maned sloth (Bradypus torquatus Illiger, 1811), endemic to the Brazilian Atlantic Forest (Miranda et al., 2023), is currently classified as Endangered (EN) due to its restricted and fragmented occupancy area and habitat, which continues to decline as a result of deforestation and anthropogenic disturbance (Santos et al., 2025). Scientific knowledge about their populations is limited (Chiarello, 2008), hindering the proposal of conservation management practices. Developing efficient survey methods to detect this threatened species is critical, as the lack of demographic data can lead to ineffective conservation planning. Santos et al. (2023) recently detected southern maned sloths (Bradypus crinitus) using thermal drones, achieving high detection rates (1.75 individuals/hour) in cold mountain forests. However, since sloths exhibit low and variable body temperatures (32 ± 5 °C; Cliffe et al., 2018; Kredel, 1928) and detection relies on the thermal contrast between the animal and the background environment (Rahman et al., 2025), the extent to which this method surpasses traditional surveys under different climatic conditions remains uncertain.
Colder conditions generally enhance thermal detection by increasing the contrast between animals and their surroundings (Beaver et al., 2020; Kays et al., 2019; Linchant et al., 2015), although this effect may reverse at lower temperature thresholds (Beranek et al., 2024). Moreover, sloths' increased use of the upper canopy for solar basking in these conditions (Montgomery and Sunquist, 1978; Urbani and Bosque, 2007) may further enhance detection by thermal drones due to reduced vegetative obstruction (Beaver et al., 2020; Kays et al., 2019). Conversely, in warmer conditions, sloths tend to stay in mid-level strata to avoid overheating (Lopes et al., 2023; Urbani and Bosque, 2007), potentially favoring ground-based methods. Given that warm lowland forests comprise the majority of the remaining habitat of the northern maned sloth (Hirsch and Chiarello, 2012), assessing the effectiveness of thermal drones in warm conditions is crucial. Moreover, if thermal drones demonstrate superior efficiency in warmer environments, they are likely to outperform traditional survey methods in cooler forests as well.
Here we aim to compare the efficiency (detection rates of individuals and occupied sites) and effectiveness (accuracy in population density estimation) of thermal drone surveys versus traditional ground-based active searches for detecting and estimating population parameters of the northern maned sloth in one of the warmest regions within its distribution. Additionally, we evaluated the cost-effectiveness associated with each method and examined the behavioral response of sloths to the approach of the thermal drone and ground-based observers as an indicator of survey invasiveness.
MethodsStudy areaThe study was conducted in seven 10.5-ha plots (324 m × 324 m) within the Atlantic Forest biome, located 2.5–4.6 km from the shoreline in the Praia do Forte district, Mata de São João, Bahia, Brazil (12 ° 34′ 39″ S, 38 ° 00′ 19″ W; Fig. 1). The plots, with over 65% forest cover, were separated by more than 500 m and confirmed to contain the target species in prior thermal drone inspections. The study area includes the Sapiranga Wildlife Refuge (REVIS Sapiranga), Camurujipe Farm, and the Quintas do Castelo neighborhood. These forests are fragmented by residential areas, streets, and highways. The vegetation is typical of lowland ombrophilous forests on coastal sandbanks, surrounded by herbaceous, shrubby, and urban ecosystems (Lopes et al., 2023). The canopy height averages 11–13 m, with emergent trees over 20 m (Mureb et al., 2023). The terrain is flat, with an elevation ranging from 10 to 30 m above sea level. The climate is tropical humid (Af, Köppen-Geiger 1918), with annual precipitation of 1250 mm and average monthly temperature ranging between 21 and 32 °C (Mureb et al., 2023; Weather Spark, 2024). During our sampling period, sunrise occurred between 5:00 and 5:20 AM, and sunset ranged between 5:40 and 5:50 PM.
Data collection, sampled plots (B), and transects within plots (C) surveyed using drones and ground-based methods for detecting maned sloths in the Praia do Forte district, Mata de São João, Bahia, Brazil.")
We conducted field sampling in October 2023 and April 2024 using two survey methods for sloth detection over three consecutive days in each plot: thermal drone surveys and ground-based active search surveys. Both methods were carried out during the same three-day period, following 10 parallel north-south transects spaced 30 m apart within each plot (Fig. 1C). Surveys followed a back-and-forth pattern, moving along one transect and returning along the adjacent one. Our survey covered a ∼30 m strip centered on each transect (15 m to each side), using such transects as spatial references to ensure complete and uniform coverage of the plots and prevent double-counting of individuals.
Thermal drone surveys were conducted by an experienced pilot and a student assistant, during early morning (5:00–8:00 AM), late afternoon (4:00–6:00 PM), and night (6:00–7:00 PM) when ambient temperatures were milder (27.8 ± 3.4 °C), avoiding peak heat periods that impair thermal detection (Fig. 2). Daily survey periods per plot were randomly assigned. We used a DJI Mavic 2 Enterprise Advanced (DJI-M2EA; DJI, Shenzhen, China) drone equipped with dual imaging systems: a thermal infrared (TIR) sensor (640 × 512 px, 30 fps, 16× digital zoom; DJI, 2021) and an RGB camera (48 M P, 32× digital zoom, with integrated 12 M P wide-angle sensor). Flight transects were preplanned using DJI Pilot software (version 2.5.1.17, DJI, 2023a), uploaded to the drone control interface, and served as navigation references.
 Visible (RGB) and (B–D) thermal infrared (TIR) images of sloths obtained with drones on sunny days at three different times: 6:00 AM (B), 8:00 AM (C), and 10:00 AM (D).")
In each survey, we manually piloted the drone along 10 transects for ∼40 min (two flights of ∼ 20 min), at predominant altitudes of ∼20 m (range: ∼20–40 m) above the canopy, maintaining an average speed of ∼4.0 m/s (range: 2.5–5.5 m/s) and with the thermal camera predominantly angled to −30° (range: −30 to −50°). We continuously adjusted the drone's altitude and gimbal in real time to account for canopy variation, maintain signal stability, and enable close-range inspection, in order to ensuring flight safety, optimize detection and maintain a consistent ∼30 m wide inspection. Although transect orientation was generally followed, we made short controlled deviations within 15 m on each side of the transect to investigate thermal anomalies or emergent tree crowns, which are often preferred by sloths (Falconi et al., 2015), promptly returning to the transect afterward.
Thermal imagery during the drone survey was monitored in real time exclusively by the pilot, using the 'white hot' palette on the integrated 5.5-inch high-brightness display (1000 cd/m²) of the DJI Smart Controller (DJI, Shenzhen, China). Upon detecting a thermal signal, the RGB camera was used to confirm whether thermal anomalies corresponded to sloths, maintaining a minimum distance of ∼15 m and limiting the observation time to a maximum of two minutes to prevent invasiveness. Real-time identification and close-range verification were necessary due to the high frequency of canopy termite mounds, which emit thermal signatures and display RGB features similar to sloths. Once a sloth was confirmed, it was photographed, and the camera was tilted to –90° (nadir) to record its GPS location. During nighttime surveys, we used the DJI M2EA Spotlight to enable visual identification with the RGB camera. Although we confirmed all detections in real-time, we filmed all flights with both sensors to allow image review in case of uncertainty in target identification. Surveys were suspended during rainfall or winds exceeding ∼10 m/s. Drone surveys totaled 13.8 h of effort across all plots.
Ground-based surveys were conducted by a separate two-person team composed of a wildlife tracker experienced in sloth detection and a student assistant, and took place during morning (8:00–11:59 AM) and afternoon (1:00–4:00 PM) hours, when ambient temperatures were higher (32.4 ± 2.3 °C) and sunlight enhanced visibility under the canopy. Whenever possible, the timing of ground-based surveys was aligned with the corresponding drone flights to ensure temporal proximity between methods. Preplanned transects used in thermal drone surveys were uploaded in a hand-held GPS and served as spatial references during the ground-based surveys. Ground-based surveys were conducted by walking on each transect at approximately 1.25 km/h, visually scanning the upper forest strata. Similar to drones, short exploratory movements of up to ∼15 m from the transect were routinely performed to inspect tall or dense tree crowns and investigate suspected targets. Upon detecting a suspected target, binoculars were used to confirm whether it was a maned sloth, and its geographical location was recorded with a hand-held GPS. The ground-based survey of a single plot took approximately 2.5 h, totaling 51.7 h for the entire sampling.
For both methods, we recorded the sampling effort (in hours), the time and location of each animal sighting, and the behavioral response of the animal during the approach. The two-person teams remained consistent for each survey method, with no exchange of information between teams during the sampling period. Given the sedentary behavior of sloths, recording GPS coordinates for each detection minimized the risk of double counting during surveys and allowed for an approximate estimation of the number of individuals per plot. Based on animal locations, the individual sloths were identified using the Euclidean distance and the temporal interval between records, as well as any external characteristics when possible. We considered detections to be of the same individual when the animal was located within 20 m on the same day, 40 m on consecutive days (one-day intervals) or 60 m on non-consecutive days (two-day intervals). These thresholds were derived from the 90th percentile of distances traveled over these intervals by eight maned sloths monitored via GPS telemetry over eight months in the REVIS Sapiranga (Giné G.A.F., unpublished data).
Efficiency and effectiveness analysisWe described the efficiency of the methods in detecting occupied sites as the percentage of plots in which each method detected the species (noting that all sites were previously confirmed as occupied, as described in the “study area” section). Additionally, following Witt et al. (2020), we measured method efficiency as the number of individuals divided by sampling effort. This value was estimated for each plot, and we compared method efficiency using a paired Student's t-test. Means ± standard deviation are provided throughout the text.
For each plot, we estimated the sloth density (Di) for each survey method (i) by dividing the number of individuals detected by the determinate method (Ni) by the sampled area (A).
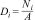
Additionally, we calculated the best density estimate for each plot, called the "naïve density" (Witt et al., 2020), dividing the number of unique individuals detected using both methods (Nb) by the sampled area (A),
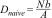
Finally, we estimated the effectiveness of each method (Ei), dividing the density estimated by the survey method (Di) by the naïve density (Dnaive) of each plot,
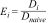
The effectiveness of the methods was compared using the Wilcoxon signed-rank test, as the normality and homoscedasticity assumptions of parametric tests were not attempted.
Cost analysisWe estimated the relative costs of the survey using two thermal drone models (the DJI-M2EA used in our surveys, and the newer DJI Mavic 3 Thermal, DJI-M3 T) and ground-based methods, categorizing expenses into six main cost groups: capital equipment, staff training, depreciation, personnel, travel, and post-processing (Table S1), based on the framework proposed by Howell et al. (2021). Capital equipment and staff training were considered upfront investments, while the other items made up the daily operational costs. We calculated the operational cost per area (ha), dividing the daily operational cost by the maximum area that can be surveyed per day following our sampling schedule, i.e., 52.5 ha (five 10.5-ha plots) for the thermal drone and 21 ha (two 10.5-ha plots) for the ground-based method. Hourly survey cost was calculated by dividing the daily operational cost by the maximum daily survey hours using our sampling schedule, i.e., five hours for the thermal drone and seven for the ground-based method. We then assessed the cost-effectiveness of each survey method by multiplying the hourly survey cost by the average time required to detect a maned sloth, based on each method's detection efficiency. All costs were estimated in Brazilian reais and converted to US dollars (R$5.79/USD, exchange rate on February 5, 2025; detailed calculations in Table S2).
Given the higher initial investment but lower operational costs of the thermal drone method, we assessed the effort required to achieve cost-effectiveness by estimating the area surveyed and number of sloth detections needed for total drone expenses (upfront + operational) to match those of traditional methods. These payback efforts were calculated by dividing the absolute difference in initial investment by the absolute difference in operational costs per area and sloth detection, respectively. The payback period was estimated by multiplying the payback efforts (per unit area and sloth detected) by the average survey time per hectare and per sloth using drones. We developed a simple linear cost evolution model to show how costs vary with sampling effort. We used the average cost of the DJI-M2EA and DJI-M3T models to evaluate thermal drone payback, since the DJI-M2EA is now discontinued. We assumed that the DJI-M3T has at least equal sampling performance and detection efficiency to the DJI-M2EA, since it is equipped with TIR and RGB sensors of the same resolution, but with enhanced zoom capacity (DJI, 2023b).
ResultsDetection efficiencyManed sloths were detected in six of seven sample plots (86% of the total) through the thermal drone survey and in three plots (43% of the total) using the ground-based survey, i.e., drones were twice more efficient in detecting sloths’ occupancy in the sampled sites. Surveying each plot with the thermal drone took 39.43 ± 7.74 min (0.65 ± 0.13 h) for a single inspection, compared to 147.62 ± 14.30 min (2.46 ± 0.24 h) using the ground-based active search method. This corresponds to a time expenditure of 3.76 ± 0.74 min (0.06 ± 0.01 h) and 14.1 ± 1.4 min (0.23 ± 0.02 h) per hectare surveyed, respectively.
During the survey, we detected 10 individuals with drones across 12 distinct detection events, while the ground survey yielded 9 individuals across 14 events. All detections and identifications were performed in real time, eliminating the need for time-consuming image review. The efficiency of sloth detection, measured by the rate of individual encounter, was significantly higher using the thermal drone (0.70 ± 0.45 individuals/hour) than the ground-based survey (0.17 ± 0.22 individuals/hour; Student t-test: t = −4.089; df = 6; p = 0.006, Fig. 3A), corresponding to a mean survey effort of 1.42 h and 5.88 h per individual encountered, respectively.
 and effectiveness (B) of thermal drone and ground-based survey methods for detecting and estimating the density of maned sloths in the Praia do Forte district, Mata de São João, Bahia, Brazil. Dashed lines connect paired samples.")
Comparison of the efficiency and effectiveness of survey methods. Comparison of the efficiency (A) and effectiveness (B) of thermal drone and ground-based survey methods for detecting and estimating the density of maned sloths in the Praia do Forte district, Mata de São João, Bahia, Brazil. Dashed lines connect paired samples.
The average naïve density of sloths was 0.20 ± 0.18 individuals/hectare. There were no significant differences between the effectiveness of the methods for estimating the sloth density (Wilcoxon test: W = 0.802; df = 6; p = 0.396, Fig. 3B). The mean estimated density using thermal drones was 0.13 ± 0.09 individuals/hectare, corresponding to a median effectiveness of 66.7% (Q1–Q3: 50–100%). The estimated density based on the ground-based survey was 0.12 ± 0.16 individuals/hectare, with a median effectiveness of 0.0% (Q1–Q3: 0.0–83.3%) due to the absence of detections in 4 out of the 7 plots. In three plots the value of density generated using the thermal drone was higher than those based on the ground-based survey, while in the other three plots, the opposite happened.
Cost analysisThe total initial investment required for sampling using a thermal drone (DJI-M2EA: $12,816.24; DJI-M3 T: $12,496.29) was, on average, 4 times higher than that of the ground-based method ($3141.55; Table S1). The daily operational costs for thermal, drones (DJI-M2EA: $399.60; DJI-M3 T: $398.37) were slightly lower compared to the ground-based method ($429.75). In contrast, thermal drone surveys yielded an operational cost per hectare (DJI-M2EA: $7.61/ha; DJI-M3 T: $7.59/ha) 2.7 times lower than the ground-based method ($20.46/ha). Similarly, the cost per individual sloth detection was 3.2 times lower with a thermal drone (DJI-M2EA: $113.49/sloth; DJI-M3 T: $113.14/sloth) than with ground-based method ($360.99/sloth). Based on our linear cost evolution model (Fig. 4), the payback point is reached after surveying approximately 740 ha (Fig. 4A). This is equivalent to 44.4 ± 7.4 h of drone survey. Alternatively, the payback point is achieved after detecting 38 sloths (Fig. 4B), equivalent to 54.5 h of drone survey. At these points, the costs of the thermal drone and ground-based survey methods equalize, and beyond the payback points, the use of drones becomes financially advantageous.
 and ground-based method (brown line)—as a function of survey effort per unit area (A) and per sloth detected (B). The payback point is reached when the lines intersect.")
Comparison of costs between thermal drones and the ground-based method. Evolution of total cost of the survey methods—thermal drone (blue line) and ground-based method (brown line)—as a function of survey effort per unit area (A) and per sloth detected (B). The payback point is reached when the lines intersect.
No noticeable behavioral response of sloths to the thermal drone’s approach was observed. In 11 of 12 detections, sloths maintained their behavior during the drone’s presence (9 resting and 2 self-grooming). In the remaining case, an individual began moving while the drone was still >60 m away, before it approached, suggesting the response was unrelated to the drone. Ground-based approaches followed a similar pattern: in all observations, individuals continued exhibiting the same behavior noted at initial detection (10 resting, 2 feeding, and 2 grooming).
DiscussionWe present the first comparative assessment of thermal drones versus traditional ground-based methods for detecting sloths and estimating population densities, evaluating efficiency, effectiveness, cost-effectiveness, and invasiveness. Our results reveal that the thermal drone method was non-invasive and significantly outperformed traditional ground-based active searches in terms of detection rates. Drones were at least three times faster in detecting individual sloths and twice as efficient in identifying occupied sites, all while operating at one-third of the cost per animal detected. This advantage stems from their ability to cover larger areas in less time – surveying 10.5-ha plots in ∼39 min compared to ∼148 min for ground-based methods – and their capacity to overcome challenges posed by the secretive behavior and habitat complexity of sloths. However, drones did not improve density estimates, suggesting their primary strength lies in rapid detection and site occupancy efficiency rather than identifying more individuals per area.
Our findings align with previous studies demonstrating the superiority of thermal drones on the traditional methods for detecting arboreal mammals, such as koalas (Howell et al., 2021; Witt et al., 2020) and primates (de Melo, 2021; Spaan et al., 2019). Despite the inherent challenges of detecting sloths due to their low and variable body temperatures, thermal drones proved at least three times more efficient than the ground-based method, reducing the average effort to detect an individual from 5.88 to 1.42 h. This remarkable improvement highlights the potential of drones to enhance the detection of arboreal mammals, particularly species like sloths, which are notoriously difficult to detect visually.
Thermal drones also showed greater success in identifying sites occupied by the target species compared to the traditional method, likely due to their consistent performance under varying habitat conditions. Although not extreme, some sampled plots posed greater challenges for ground-based surveys because of topographic irregularities, tall canopies, and dense understory vegetation, which restricted observer movement and limited visibility of the canopy. These same structural features can also hinder aerial thermal detection; however, as sloths predominantly occupy the outer canopy (Lopes et al., 2023), such obstacles likely impair ground-based detection more severely than they affect drone-based surveys. The consistent performance of drones, combined with their ability to access remote or hard-to-reach locations (Anderson and Gaston, 2013) and rapidly scan large forest areas (Linchant et al., 2015; Witt et al., 2020), makes them a promising tool for habitat occupancy and population abundance studies accounting for imperfect detection (Ryan et al., 2025), which require repeated sampling across time and/or space (MacKenzie et al., 2002). Drone technology can streamline data collection for such designs, significantly reducing human effort while maintaining high efficiency (Howell et al., 2021; Linchant et al., 2015; Witt et al., 2020). These properties can improve the feasibility of such studies, potentially enabling significant advances in our understanding of the distribution of arboreal species and their response to disturbances (Beranek et al., 2024; Howell et al., 2021; Ryan et al., 2025).
Interestingly, thermal drone and ground-based surveys showed similar effectiveness in generating density estimates, contrasting with previous findings on koalas (Witt et al., 2020), but aligning with results for orangutans (Rahman et al., 2025). While thermal drones detected sloths in more sites (plots) and at a faster rate, the number of sloths per unit area was similar between methods, at least when the same number of repeated surveys (n = 3) was conducted. At the plot scale, vegetation obstructions present challenges for both approaches, as arboreal animals may evade thermal drones when occupying lower forest strata, including the mid-canopy (Witt et al., 2020), and remain undetected by ground-based surveys in the upper canopy. This may be especially significant in dense tropical forests (Haysom et al., 2021), such as those surveyed in this study. Our results indicate that individuals detected by drones were often missed by the traditional method, and vice versa, emphasizing that these are complementary approaches. This complementarity is further reinforced by the differing optimal survey times. Drones tend be more effective during cooler periods with lower solar radiation, which enhanced thermal contrast (Beaver et al., 2020; Rahman et al., 2020, 2025), whereas diurnal ground-based active searches depend on solar radiation for improve the visibility under canopy. In any case, it is worth noting that repeated surveys may improve the accuracy of both methods, potentially achievable with less effort and cost using thermal drones compared to the traditional ground-based method. Further research is needed to clarify the effect of the number of repeated surveys on density estimates for both methods.
We compared the cost-effectiveness of the two methods, as this is a critical factor in decision-making when designing biodiversity studies or monitoring programs (Beranek et al., 2024). Our findings highlight a clear trade-off between initial investment and long-term cost efficiency. While the initial cost of thermal drones is higher, the operational cost per survey area and detected animal is significantly lower, equalizing the costs after a relatively medium time (∼740 ha surveyed or ∼38 sloths, corresponding to ∼44 and 55 h of drone survey, respectively). Similar findings were reported by Howell et al. (2021), who compared the costs of drone surveys with other two traditional methods for koala detection, a species ecologically comparable to sloths. This demonstrates that for these species, the drones are an excellent option for medium- to long-term studies and medium- to large-scale surveys. In addition, the upfront cost of thermal drones is expected to decrease with technological advancements (Rahman et al., 2025), as seen between the DJI-M3T and DJI-M2EA. The rapid evolution of drone technology, including higher-resolution sensors and enhanced zoom capabilities (Church et al., 2024), standardized flight protocols, and other methodological advances, promises even greater accuracy in population estimates (Beaver et al., 2020; Hodgson et al., 2017; Rahman et al., 2025; Ryan et al., 2025).
While thermal drone surveys offer key advantages for detecting arboreal mammals (Howell et al., 2021; Spaan et al., 2019; Witt et al., 2020), they also have important limitations. First, the detection efficiency is species-dependent and some arboreal mammal species are not efficiently detected by thermal drones (Rahman and Rahman, 2021). Second, time and cost per animal detected may vary depending on species density, forest structure, topography, climatic conditions, and the specific technology and methodology adopted, thus making the payback period inherently context-dependent (Howell et al., 2021). Beyond high equipment costs, the need for elevated launch sites and trained field operators may reduce the cost-effectiveness of thermal drones compared to remote monitoring systems, especially when the objective is to detect presence–absence rather than obtain count data (Beranek et al., 2024; Howell et al., 2021). In this scenario, our findings should not be interpreted as a generalized endorsement of thermal drones for all applications and contexts, but rather as evidence of their relative performance compared to the traditional ground-based method under controlled, standardized conditions, providing practical insights to support future monitoring efforts focused on sloths
We demonstrated that thermal drones significantly outperform traditional ground-based active searches in both efficiency and cost-effectiveness for sloth detection, allowing non-invasive acquisition of both occupancy and count data, aligning with findings for other arboreal mammal species (de Melo, 2021; Howell et al., 2021; Kays et al., 2019; Spaan et al., 2019; Witt et al., 2020). This approach can facilitate the identification of critical habitats and species responses to environmental change, enabling timely conservation and mitigation actions. Additionally, the method’s capacity to rapidly detect and accurately locate individuals highlights its potential for pre-deforestation rescue operations to minimize sloth mortality during forest clearing. Integrating thermal drones into monitoring protocols can greatly enhance our ability to study and protect arboreal mammals such as sloths, leading to more effective and targeted conservation outcomes.
FundingThis work was supported by the Universidade Estadual de Santa Cruz (UESC, PROPP: 073.6764.2024.0003434-74) and the Financiadora de Estudos e Projetos (FINEP). Additionally, the Postgraduate Program in Ecology and Biodiversity Conservation (PPGECB) at UESC provided essential materials for conducting this research, and the Fundação de Amparo à Pesquisa da Bahia (FAPESB) awarded a fellowship to G.N. dos Santos. This work was also supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior–Brazil (CAPES) (Finance Code 001) through a Doctoral Scholarship granted to G.A.-F.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We thank the logistical support provided by the Aruá Wildlife Observation and Applied Ecology & Conservation Lab. We thank the authorizations provided by the Instituto Chico Mendes de Conservação da Biodiversidade (SISBIO 94705-1), Ethic Committee from UESC (CEUA 038/24), managers of the Refugio de Vida Silvestre Sapiranga and local residents of the study area.
The following is Supplementary data to this article:

