







Continuous systematic monitoring is necessary to measure the success or failure of mineland rehabilitation. Here, we aimed to outline how measures of phylogenetic diversity and phylogenetic community structure complement existing monitoring frameworks along a waste pile rehabilitation chronosequence in Carajás National Forest, Brazil, including primary rainforest as the desired outcome. We pruned the megatree R20160415.new to tree species found in this study and computed the phylogenetic diversity (PD), mean pairwise distance (MPD) among taxa and individuals and their standard effect sizes (ses.PD and ses.MPD) via the 'phylogeny pool' null model. PD and MPD increased with rehabilitation progress. Dominance of Fabales species caused phylogenetic clustering during early rehabilitation stages. This clustering disappeared in older stands and switched to phylogenetic overdispersion at reference sites, indicating that mechanisms impeding the recruitment of species from certain phylogenetic lineages during initial stages, such as environmental filtering, tend to decrease with rehabilitation advancement. Furthermore, this pattern indicates the recovery of coexistence-stabilizing interactions with rehabilitation time, even if a constrained number of species was used to launch rehabilitation. Positive associations between all phylogenetic variables and previously determined measures of environmental quality indicate the importance of community assembly processes and niche diversification for ecosystem performance in rehabilitating minelands.
Mineland rehabilitation aims to restitute biodiversity and ecosystem functioning as closely as possible to premining levels (Gastauer et al., 2018a), representing an important pillar of the mitigation hierarchy (World Economic Forum, 2016) for reconciling economic development with the conservation of biodiversity and ecosystem services (Lovejoy and Nobre, 2019). When implemented successfully, such rehabilitation promotes self-perpetuating communities and ecosystems (Gann et al., 2019) and becomes an integral part of conservation planning and implementation (Volis, 2019). Continuous systematic monitoring of ecosystem rehabilitation is necessary to detect changes in plant communities along rehabilitation trajectories, measure the success or failure of activities and provide important information about the return of ecosystem functions and biodiversity after mining (Gwenzi, 2021).
Traditional environmental monitoring programs survey field variables related to species composition, abundance, and taxonomic diversity as well as vegetation structure and key ecosystem functions such as carbon sequestration to track the trajectory of rehabilitating minelands (Gann et al., 2019). Methods for integrating different environmental properties into a measure of rehabilitation success are available (Gastauer et al., 2021, see methods for details), and phylogenetic ecology, which links evolutionary history and short-term ecological processes, may provide additional tools for the assessment of rehabilitating minelands (Gastauer et al., 2018b; Hipp et al., 2015). This is because phylogenetic diversity (PD) can be a useful proxy for high-dimensional functional trait diversity (Tucker et al., 2018), maximizing communities' abilities to resist and/or recover from disturbances (Augeraud-Véron et al., 2019). Thus, the lack of phylogenetic groups in rehabilitating minelands may lead to ecological function decay, hinder the return of ecosystem services, and reduce community resistance and resilience (Maza-Villalobos et al., 2011).
Specific traits and adaptations may be required for plant recruitment in degraded minelands, commonly characterized by low nutrient availability, a lack of mutualism, high temperatures and elevated radiation (Mukhopadhyay et al., 2016). If such adaptations occur in only a few evolutionary lineages, such environmental filtering may cause phylogenetic clustering, i.e., the coexistence of closely related species within the community (Ginocchio et al., 2017). In contrast, interspecific interactions such as competition or facilitation limit niche overlap between coexisting species (Carrión et al., 2017). When ecological niches are conserved within evolutionary lineages, such that close relatives tend to resemble each other, the phylogenetic distance between coexisting species tends to increase when interaction-mediated coexistence is relevant for community assembly (Godoy et al., 2014).
Because environmental constraints of degraded minelands are expected to be alleviated with the establishment of vegetation cover and the return of soil function (Gastauer et al., 2018a,b,c), the recovery of plant–plant interactions should shift the phylogenetic structure of plant communities from clustering to overdispersion (Swenson and Enquist, 2009). The return of interactions is furthermore expected to contribute to the success, stability, resistance, and resilience of the rehabilitating areas (Lake, 2013). Accordingly, the evaluation of phylogenetic relatedness in plant communities along rehabilitation trajectories may be important, especially when we aim to infer community assembly along rehabilitation chronosequences. In such efforts, the integration of phylogenetic indices into monitoring frameworks is paramount.
Here, we present an analysis of phylogenetic structure and diversity along a rehabilitating mineland chronosequence in the Carajás National Forest, eastern Amazon, Brazil. This region harbors the world’s most important reserves of iron ore, and comprehensive open-cast mining requires large-scale mineland rehabilitation. The objective was to evaluate these features and provide feedback to enhance the rehabilitation processes, understand the underlying processes of community assembly and outline how phylogenetic measures complement existing monitoring frameworks. We expected that (i) tree communities in initial rehabilitation stages would be dominated by only a few phylogenetic clades, while clades would be more evenly distributed in older stages and at reference sites (random or overdispersed); (ii) phylogenetic clustering would decrease with the advancement of rehabilitation, such that phylogenetic structure would become more similar to reference values from natural ecosystems over time; and (iii) metrics of diversity and phylogenetic structure would be related to the success of rehabilitation activities.
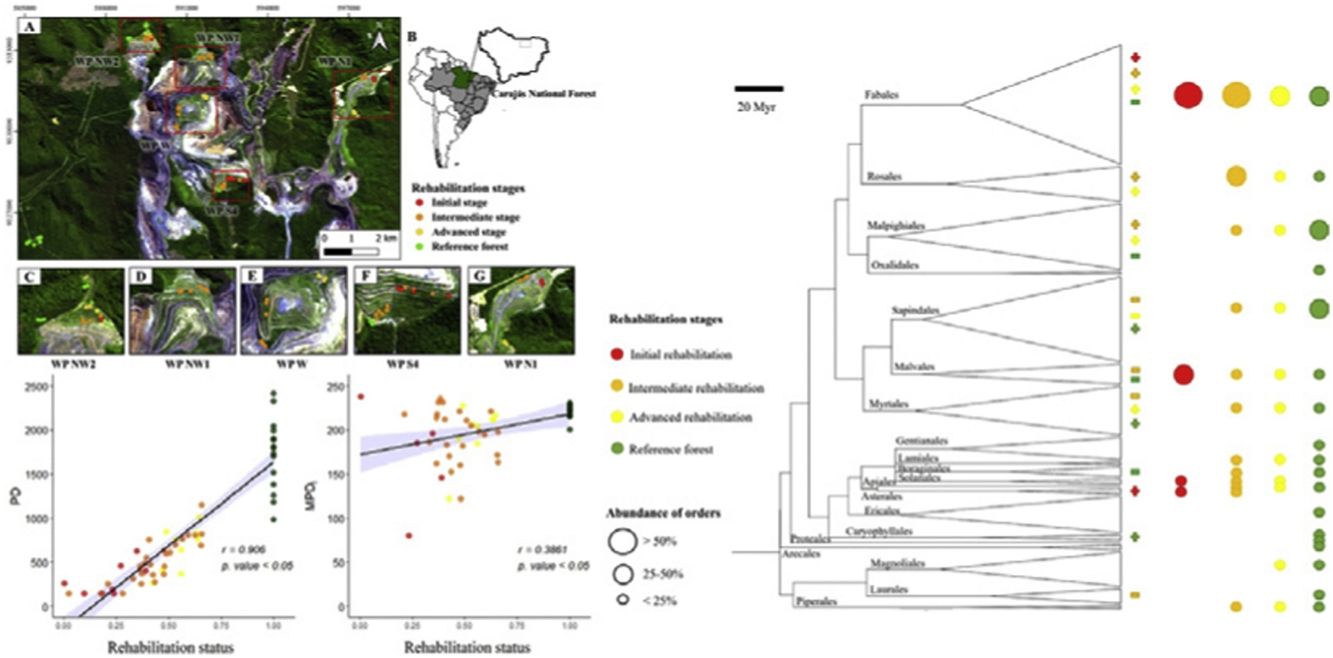
MethodsStudy siteThis study involved five rehabilitating waste piles (WPs: S4, NW1, NW2, N1 and West) from the open-cast N4-N5 iron mining complex within the Carajás National Forest, eastern Amazon, Brazil (Fig. 1). The climate of the region is classified as Aw according to the Koppen classification (Alvares et al., 2013), i.e., a tropical warm climate with dry winters and rainy summers. The average annual temperature ranges between 21 °C and 22 °C, and the mean annual precipitation is between 1900 and 2000 mm.
For the mining of high-grade iron ore, the removal of vegetation and overburden is necessary. Overburden is deposited at nearby sites, forming large WPs structured by distinct benches that are rehabilitated when the filling of one level is completed. Benches are approximately 60 m in height, with slope angles varying between 40 and 45°, and soils are sandy clays with intermediate nutrient availability compared to that of surrounding forests (Guedes et al., 2021, Supplementary Material 1). For rehabilitation, WP benches are hydroseeded. Specifically, fertilizers (600 kg ha−1 NPK 4-14-08 and 10 kg ha−1 micronutrients via FRITAS BR12) and seeds of fast-growing, exotic nitrogen-fixing legumes and grasses (250 kg ha−1) were mixed with water and sprayed on the slope surface (Supplementary Material 2). Organic mulches (2 Mg ha−1) were added to guarantee the fixation of seeds and fertilizers. To trigger natural succession toward reference ecosystems, the mixture was enriched with seeds from native species from surrounding ecosystems (50 kg ha−1). The number and identity of seeded species depends on seed availability during the planting period (from November to March) and varies slightly from year to year. As mining and mine waste deposition are ongoing activities in the N4-N5 complex, individual benches of the examined WPs comprise different rehabilitation stages, with older rehabilitation stages generally being situated in the lower parts of the piles, while younger stages are found toward their tops.
SamplingBefore fieldwork, we examined the company's reports, maps, and satellite images to identify benches of different rehabilitation ages in the five WPs. Between February and April 2019, three plots (10 × 20 m) were installed in each of the previously identified sections (based on rehabilitation age). Plots were installed at the intermediate portions of the benches to minimize the risk that the plots’ substrates would be disproportionately affected by erosion or sedimentation of finer soil particles. Furthermore, three plots were sampled in each of five distinct reference sites covered by dense evergreen forest, totaling 60 plots. We selected this type of reference ecosystem because these forests correspond to the predisturbance ecosystems and are declared rehabilitation target.
Within plots, all trees with diameters at breast height ≥3 cm were tagged and identified to the species level. Species not recognized in the field were collected and identified by herbarium comparison and specialists. Species classification followed that of APG IV (The Angiosperm Phylogeny Group, 2016), and nomenclature was used as proposed by tropicos.org.
According to rehabilitation age, we grouped the rehabilitating benches into (i) initial (revegetation dates up to three years prior, 3 plots each in WPs NW2, S4 and N1), (ii) intermediate (4-6 years, 3 plots each in WPs NW2 and N1 and 6 plots each in WPs NW1, W, S4) and (iii) advanced (vegetation 7 or more years old, 3 plots each in WPs NW2, W, S4 and N1) rehabilitation stages. These thresholds were defined on the basis of shrub encroachment being observed three years after the beginning of rehabilitation activities and canopy closure occurring after seven years. Together with reference forests (6 plots near WP NW2 and at a site called Arenito, 3 plots near WP S4), we sampled a four-stage rehabilitation chronosequence. Due to irregular waste deposition and subsequent rehabilitation as a function of mining advancement, the rehabilitation stages were not equally represented within monitored WPs, resulting in an unbalanced sampling design.
Phylogenetic diversity and structureFor phylogenetic tree construction, the megatree R20160415.new containing all angiosperm families was pruned to include only the tree species recorded in this study using the ‘phylomatic’ function of the Phylocom-4.2 program (Webb and Donoghue, 2005). The resulting tree was calibrated ('bladj' function, Phylocom-4.2, Webb et al., 2002) so that the branch lengths corresponded to the evolutionary distances between species. To do so, age estimates for internal nodes proposed by Magallón et al. (2015) were used.
PD was computed as the sum of branch lengths connecting species from each plot (Faith 1992). Additionally, using the ‘picante’ package in R (R Development Core Team, 2018), we computed the mean pairwise PD (MPD) among distinct species (MPDt) and individuals (MPDi) from each plot (Webb et al., 2002; Fine and Kembel, 2011). Small MPD values indicate communities assembled by closely related species, and large MPD values occur when coexisting species are separated by deep evolutionary splits. While MPDt is based on species presence and allows comparisons of community structure considering the influence of rare species, the abundance-weighted analysis (MPDi) aims to reveal the influence of dominant species on community structure. For the latter metric, phylogenetic distances among individuals from the same species were not considered for computation.
To test whether coexisting species from each rehabilitation stage were more (phylogenetic clustering) or less (phylogenetic overdispersion) related than expected by chance, phylogenetic structure for each plot was computed as the standard effect sizes [(observed values – mean of expected values)/standard deviation of expected values] of all phylogenetic metrics described above. Expected means and standard deviations were derived from a null model randomizing community composition 10,000 times (Gastauer et al., 2017). Positive values (observed > expected) indicate phylogenetic overdispersion, and negative values (observed < expected) indicate phylogenetic clustering. Within the null model, the species richness (and abundance distribution for abundance-weighted indices) of each plot was kept constant, and all species from the phylogeny had the same probability of joining the randomized community. As all species recorded in this study were considered equally able to colonize any plot, we tested how factors related to rehabilitation age influence phylogenetic community composition (Gotelli and Graves, 1996).
To test whether the phylogenetic structures of rehabilitation stages differed from random expectations, Wilcoxon rank tests against 0 were performed. Marginal phylogenetic structure was considered when p values varied between 0.1 and 0.05. To address the unbalanced sampling design (see Sampling Section for details), Kruskal–Wallis and post hoc Dunn tests were performed to test for differences in phylogenetic structure and PD between rehabilitation stages.
Finally, we applied the function ‘nodesig’ of Phylocom-4.2 to check for lineages represented by more or fewer descendants than expected by chance within distinct rehabilitation stages (Webb et al., 2002). As a null model, we randomly sampled species occurrences among rehabilitation stages.
The main criticisms of the megatree approach applied here are the emergence of polytomies in species-rich families due to a lack of phylogenetic resolution (Gastauer et al., 2018c; Swenson, 2009). To determine how inadequate phylogenetic resolution influences the outcomes for PD and phylogenetic structure, we randomized all polytomies 999 times using the 'multi2di' function of the 'picante' package, dated the fully resolved phylogeny as described above and computed all phylogenetic metrics. Outcomes from unresolved and randomly resolved phylogenies were compared using linear correlations. Because we found strong correlations between different phylogenetic metrics computed from unresolved and resolved phylogenies (R2 and mean z values larger than 0.9), we concluded that inadequate phylogenetic resolution only marginally influenced our results (Supplementary Material 4), although statistical inference of further sources of phylogenetic resolution, e.g., the hypothetical nature of the phylogenetic tree or missing extant taxa, may be necessary (Rangel et al., 2015).
Rehabilitation statusTo check whether phylogenetic metrics were related to environmental quality, we computed rehabilitation status as described by (Gastauer et al., 2020) using field-surveyed environmental variables related to vegetation structure (tree density, basal area, and leaf area index), community diversity and composition (Shannon index of tree diversity, percentage of native trees and percentage of nonnative trees) and ecological processes (soil organic matter, functional richness and aboveground biomass). For methodological details on the survey of these variables, see Supplementary Material 5. In brief, rehabilitation status corresponds to the portion of environmental success already achieved by the rehabilitation trajectory considering the overall rehabilitation trajectory necessary to transform degraded minelands into ecosystems with levels of vegetation structure, diversity and ecological processes similar to those of the reference ecosystems. Targeting the same WPs from the Carajás National Forest, Gastauer et al. (2021) used 27 environmental variables but showed that a much smaller set was able to explain environmental status reliably; additionally, the authors identified the Shannon index of tree diversity as the best predictor of overall environmental status at the study site.
To measure relationships between phylogenetic metrics and rehabilitation status, we computed Spearman’s rank correlation coefficient to address the unbalanced sampling design. For this, we used the ‘cor.test’ function of the R ‘stats’ package. We constructed illustrative plots with ‘ggscatter’.
ResultsIn total, 1536 trees of 232 species and 21 orders were recorded. With 37 species, Fabales was the most species-rich order (Fig. 2). With 179 species from 18 orders, the species richness at the reference sites was higher than that at the rehabilitation sites (total of 67 species, Supplementary Material 1). Among the rehabilitation sites, the advanced stage included 49 species and 11 orders, followed by the intermediate stage with 43 species in 12 orders and the initial stage with 7 species and 4 orders.
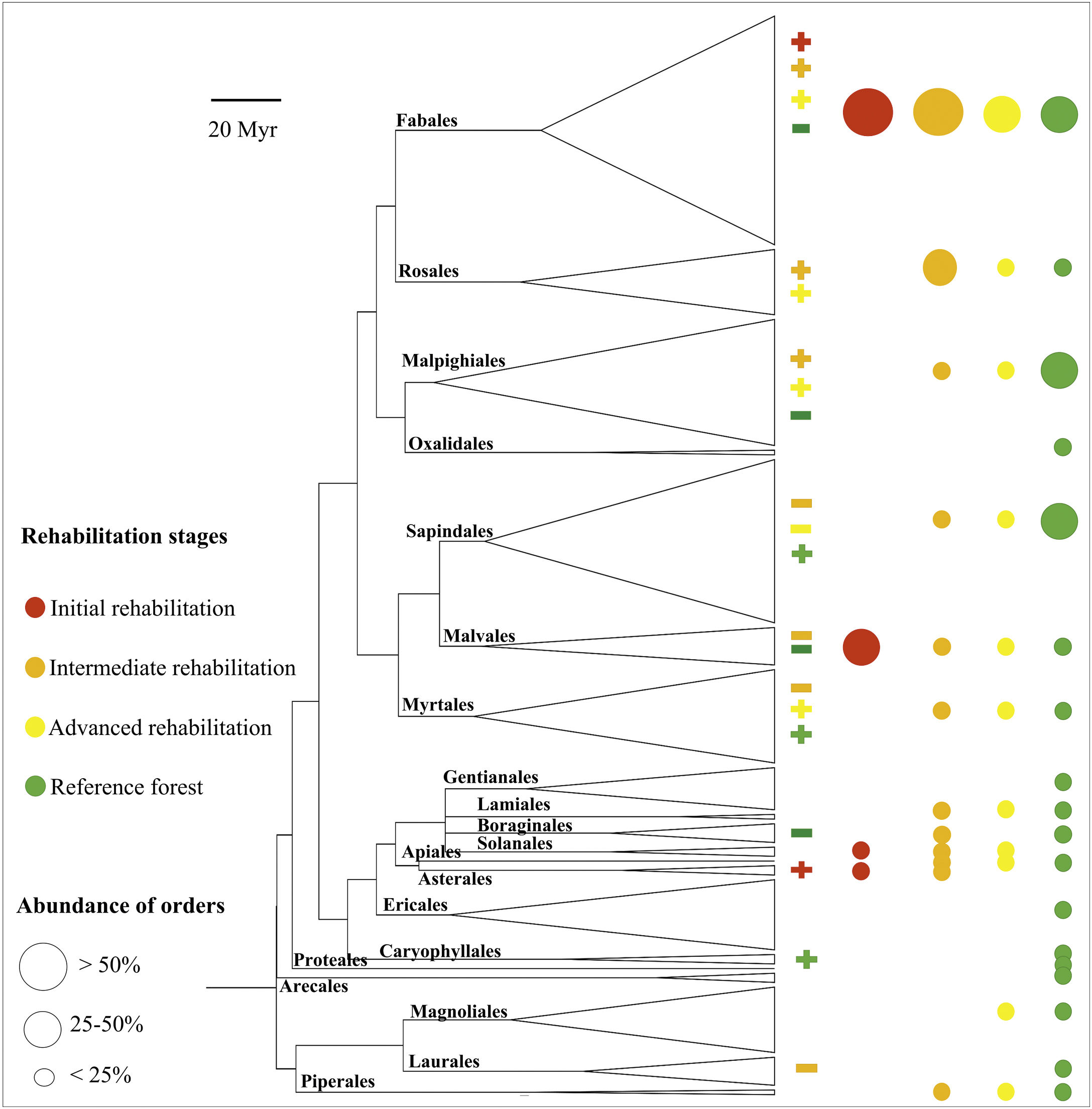
Phylogeny of angiosperm tree species sampled in this study. Positive and negative signals indicate more or fewer daughter taxa than expected by chance, respectively, from nodesig analysis within distinct rehabilitation stages (p value ≤ 0.05; for details on nodesig analysis, see Supplementary Material 3).
All orders except Solanales and Asterales were detected at the reference sites, where more descendants than expected by chance were detected in the orders Sapindales, Myrtales and Caryophyllales (Fig. 2), indicating a lack of these orders in rehabilitation stages. Trees of only four orders were found in the initial rehabilitation stages, including Fabales and Asterales, with more descendants than expected by chance. Oxalidales, Gentianales, Ericales, Caryophyllales, Proteales, Arecales (palms) and Laurales were lacking in the intermediate and advanced rehabilitation stages, where Fabales, Rosales, Malpighiales and Myrtales were represented by more species than expected by chance, indicating their importance for phylogenetic clustering. The abundance of angiosperm orders varied considerably among and within the rehabilitation stages (Fig. 2).
Reference areas showed higher PD than rehabilitating sites (Fig. 3), and increases in PD from initial to intermediate and advanced stages were detected. MPDt and MPDi values were higher at reference sites than at rehabilitating sites, but no differences between rehabilitation stages were found. Regarding phylogenetic structure, marginal overdispersion (0.1 < p value < 0.05) was found in reference forests (Fig. 3). In the initial stage, ses.MPDi indicated phylogenetic clustering, and ses.PD and ses.MPDt indicated marginal phylogenetic clustering. The low significance level is most likely due to the small number of plots with two or more shrub species necessary to compute these phylogenetic metrics (Supplementary Material 1). ses.PD and ses.MPDt indicated phylogenetic clustering in the intermediate and advanced stages, whereas phylogenetic structure measured by ses.MPDi in the advanced stage did not differ from the null expectation. ses.PD and ses.MPDt values were higher in the reference forests than in the rehabilitation stages. When measured by ses.MPDi, advanced stages did not differ from reference sites or from initial or intermediate rehabilitation stages (Fig. 3). All the metrics of phylogenetic structure and PD were positively associated with rehabilitation status, and correlation coefficients were higher for PD and MPDi than for MPDt or the standard effect transformations of the metrics (Fig. 4).
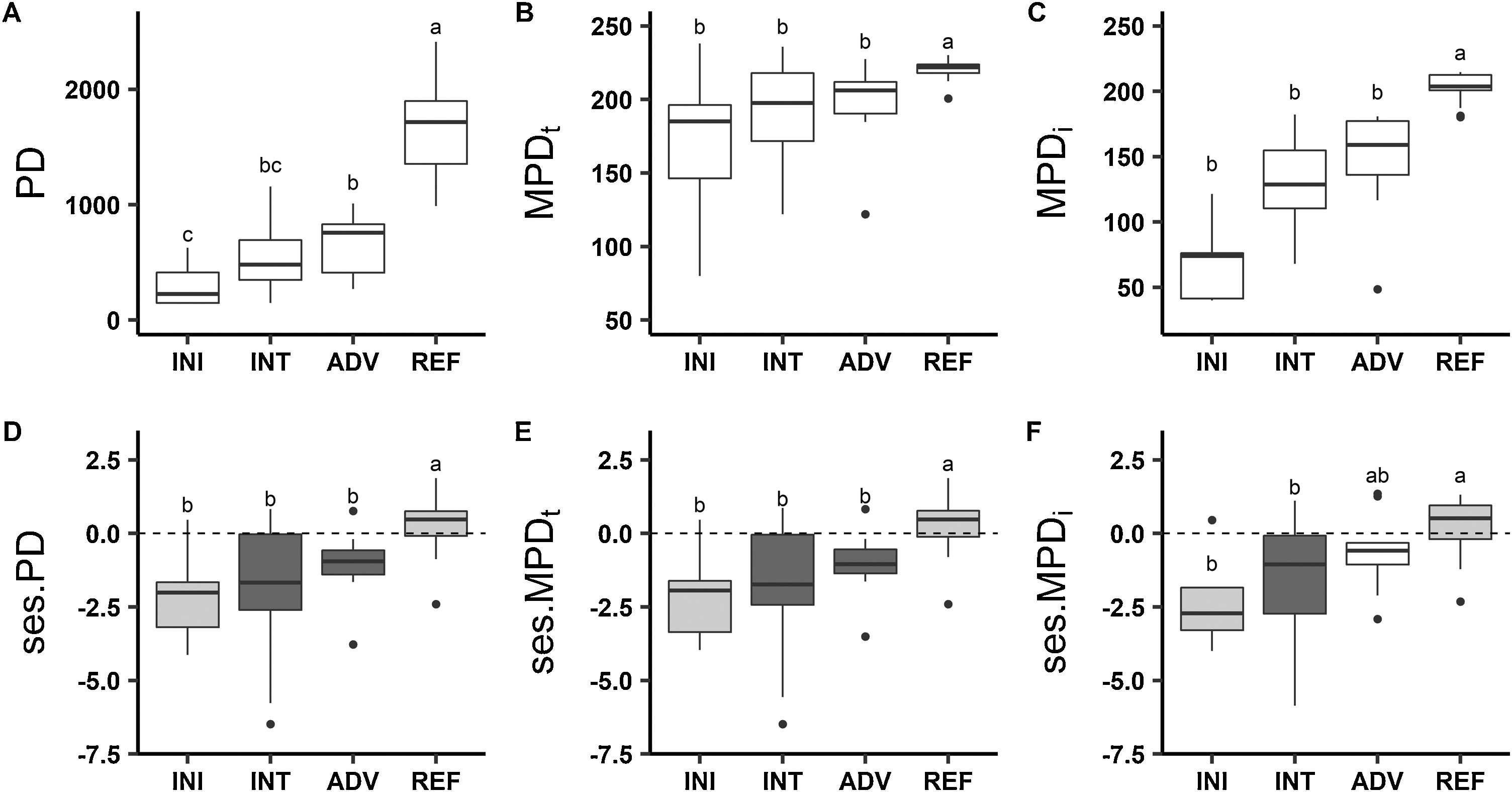
Phylogenetic diversity (A. PD - phylogenetic diversity, B. MPDt - mean pairwise phylogenetic distance among taxa and C. MPDi - mean pairwise phylogenetic distance) and structure (D. ses.PD - standard effect size of phylogenetic diversity, E. ses.MPDt - standard effect size of mean pairwise distance among taxa, F. ses.MPDi - standard effect size of mean pairwise distance) comparisons among initial, intermediate and advanced rehabilitation stages and reference sites. Different letters indicate significant differences between sites (p ≤ 0.05). Darkly shaded bars indicate phylogenetic structures differing from the null expectation (p ≤ 0.05), and lightly shaded bars differ marginally from the null expectation (0.1 ≤ p < 0.05).
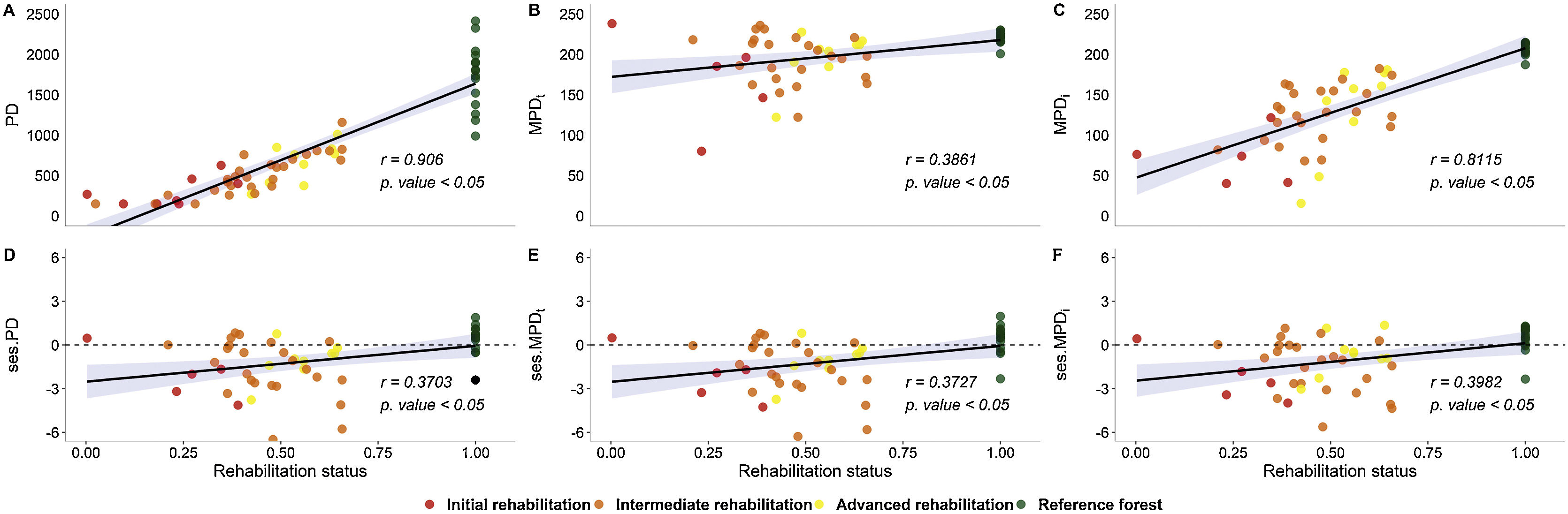
Rehabilitation status in comparison to phylogenetic diversity and structure (A. PD - phylogenetic diversity; B. MPDt - mean pairwise phylogenetic distance among taxa; C. MPDi - MPD among individuals; D. ses.PD - standard effect size of PD; E. ses.MPDt - standard effect size of MPD among taxa; F. ses.MPDi - standard effect size of MPD among individuals).
As expected, we observed shifts in tree community composition and phylogenetic metrics along the analyzed rehabilitation chronosequence. First, Fabales and Malvales dominated tree communities beginning in the initial rehabilitation stage, and further clades established with rehabilitation advancement. Second, (marginal) phylogenetic clustering (0.05 < p values ≤ 0.1) in early rehabilitation stages indicated the presence of mechanisms that limit the establishment of species belonging to a few evolutionary clades. With time, all phylogenetic metrics used here increased and became more similar to values found at reference sites, indicating that actual rehabilitation efforts met desired trajectories (Gastauer et al., 2021). If this trend continues, rehabilitating minelands may indeed become habitats for species from distinct phylogenetic lineages. Third, phylogenetic metrics were positively associated with the environmental quality of the rehabilitation sites and should be further explored to outline the importance of evolutionary history for vegetation structure, community diversity and ecological processes.
More daughter lineages from the Fabales order than expected by chance caused phylogenetic clustering in early rehabilitation stages. Due to their adaptations to high-radiation and nitrogen- and water-limited soils (Gei et al., 2018), higher seed viability and higher growth rates than those of seeds from other families (Ramos et al., 2019), nitrogen-fixing legume species perform better in these environments than other families and are recommended for further mineland rehabilitation purposes (Jia et al., 2020). Following these recommendations, a large portion of seeds of nitrogen-fixing species from the Fabales clade were applied during the hydroseeding procedure (see Supplementary Material 2 for details), contributing to the observed dominance, on the one hand.
On the other hand, similar patterns, i.e., phylogenetic clustering (Satdichanh et al., 2019; Yu et al., 2019) and high abundances of species from the Fabaceae family (Maza-Villalobos et al., 2011), have been reported for early rehabilitation stages in the literature, including in naturally regenerating ecosystems, and have been associated with environmental filtering. Thus, the observed phylogenetic clustering is also consistent with the presence of environmental filters that select for species with specific traits (Gastauer et al., 2018b). Increases in MPD values and shifts toward phylogenetic randomness, as detected by abundance-weighted measures of phylogenetic structure (ses.MPDi), within the rehabilitation chronosequence indicate the colonization of the sites by distantly related clades. In addition to the colonization by evolutionary lineages new to the sites, larger increases in abundance-weighted MPDi than in MPDt values (based on presence-abundance data only) with rehabilitation advancement highlight the above-average recruitment of lineages that were not highly represented during the initial stage.
Although the colonization of distantly related species with rehabilitation advancement may result from longer growth periods of hydroseeded species from further clades to achieve inclusion criteria, this indicates that eventual environmental filters during early stages are rapidly alleviated with rehabilitation advancement. Thus, mineland colonization may indeed occur randomly or uniformly (and not in a clustered manner) throughout the phylogenetic tree, and diverse plant communities containing representatives of distinct evolutionary clades can rapidly establish. The observed switch from phylogenetically clustered to phylogenetically random or overdispersed communities over time further indicates the emergence of interaction-mediated coexistence in the advanced rehabilitation stage (Martínez-Ramos et al., 2021). This is because plant–plant interactions such as competition or facilitation tend to limit the phylogenetic similarity among coexisting species when ecological niches are conserved within evolutionary lineages (Webb et al., 2002), contributing to niche diversification with rehabilitation advancement (Cadotte et al., 2016).
Correlations between phylogenetic metrics and overall rehabilitation status demonstrated partial recovery of phylogenetic community structure and PD simultaneously with improvements in environmental quality. Considering shifts from phylogenetic clustering to overdispersion with rehabilitation advancement, consistent with the alleviation of environmental effects and the emergence of coexistence-stabilizing plant–plant interactions, our data highlight the importance of niche diversification for the ecosystem performance of rehabilitating minelands (Enquist et al., 2015), although the underlying mechanisms require further investigation.
The findings of this study have two major implications. First, the correlations between phylogenetic metrics and overall environmental quality show that phylogenetic structure and PD recover during the rehabilitation process together with further ecosystem properties. Thus, mineland rehabilitation may indeed contribute to mining impact mitigation, as these habitats may become home to a wide range of species from distinct evolutionary clades and a considerable portion of PD. Second, mechanisms restricting the colonization and establishment of phylogenetically distinct taxa in early mineland rehabilitation stages may hinder the enhancement of tree diversity found to be paramount in maximizing rehabilitation success (Gastauer et al., 2021). Therefore, using a narrow set of species to initiate the rehabilitation process may alleviate the environmental filters and over time allow a more diverse set of species to colonize. When needed, enrichment planting of a phylogenetically diverse set of species at later stages can help enhance diversity and increase rehabilitation success. Candidate species for enrichment plantings may be found in the orders Oxalidales, Ericales, Caryophyllales, Proteales and Arecales, which were found in the reference forests but not at the rehabilitation sites.
ConclusionOur study confirms previous findings regarding rehabilitating chronosequences, such as the convergence of important ecosystem characteristics toward those of reference sites. Eventual environmental filters or further mechanisms hindering the establishment of some lineages in the initial stages of mineland rehabilitation tend to be overcome with time, although important ecosystem properties such as PD are not completely recovered within the observed periods. Strong associations between the overall environmental quality of rehabilitating minelands and phylogenetic indicators highlight the importance of niche diversification in the ecosystem performance of rehabilitating minelands.
Conflict of interestNone declared.
This work was funded by Instituto Tecnológico Vale (RAD I and Pará Resident Program) and additional support provided by CNPq to SJR (305831/2016-0).
The following is Supplementary data to this article:


