




Mesocarnivores are sensitive to global environmental changes, thereby serving as valuable sentinel species for assessing the ecological effects of anthropogenic disturbances. In human-modified landscapes, understanding mesocarnivore community structure and species habitat use is useful for guiding conservation strategies and preserving key ecosystem functions and services. Using camera-trap data, we evaluated the occupancy and detection probabilities of mesocarnivore assemblages in two protected areas of the Atlantic Forest, northeastern Brazil: RPPN Estação Veracel (sustainable-use) and Pau Brasil National Park (strictly protected). Ten mesocarnivore species were recorded, with higher species richness and occupancy rates in the strictly protected area. Occupancy and detection probabilities varied substantially among species and over time, with no strong responses to site-scale covariates. Nasua nasua was favored in sites with larger tree basal area, while Cerdocyon thous avoided these sites. Didelphis aurita was more common in sites distant from roads, contrasting with C. thous. All species, particularly Leopardus sp. and N. nasua, tended to use areas near reserve boundaries. Despite the high species diversity, low average occupancy suggests barriers limiting population growth, with some species benefiting from anthropogenic disturbance, underscoring the complexity of mesocarnivore roles in tropical ecosystems.

Human activities have substantially modified natural ecosystems (Dirzo et al., 2014), leading to ecological consequences, such as changes in resource availability, species distribution, and interspecific interactions (Liu et al., 2007). In many cases, anthropogenic influence can exert a more pronounced effect on wildlife populations (e.g. changes in distribution or abundance; Wolff et al., 2019; Newbold et al., 2020) and communities (e.g. changes in species richness, evenness, and trophic structure; Roemer et al., 2009; Magioli et al., 2019) than the natural environmental conditions (Nagy-reis et al., 2017).
Wild mammals have been severely affected by human activities, primarily through overexploitation and habitat loss and degradation, which pose significant threats to their survival (Galetti et al., 2021). These impacts are particularly evident in tropical forests (Dirzo et al., 2014) such as the Atlantic Forest, a biodiversity hotspot (Ribeiro et al., 2011). In this biome, most of the remaining fragments are highly defaunated, with declining populations of medium- and large-sized mammals (Bogoni et al., 2018). This context may exacerbate shifts in species dominance and alter trophic cascades within forest fragments (Roemer et al., 2009), with distinct disturbance regimes driving different tendencies in the species assemblages. For instance, the extirpation of top predators aligned with increasing food supply in agricultural fields may drive an increase in large herbivores abundance, consequently impacting plant assemblages in forest fragments; but overhunting and habitat loss may restrain this group, leading to the ecological release of smaller herbivores, usually seed predators (Pires and Galetti, 2023).
Mesocarnivores – predators weighing less than 15 kg and occupying an intermediate position in the trophic web (Roemer et al., 2009) – are another functional group of medium-sized mammal that may benefit from top predator extirpation and from resource surplus in anthropogenic matrices (Pires and Galetti, 2023). These species contribute to key ecological functions, including predation of invertebrates and small vertebrates, and primary and secondary seed dispersal (Cazetta and Galetti, 2009). Although mesocarnivores are considered resilient and expected to increase in abundance with the removal of larger competitors (Estes et al., 2011), 91% of small carnivorans are threatened by habitat loss – particularly from agricultural expansion (85%) – as well as by vehicle collisions and retaliatory hunting throughout their distribution (Marneweck et al., 2021). Human disturbances can have both negative and positive impacts on mesocarnivores. Changes in the distribution and abundance of mesocarnivore population can serve as indicators of environmental change, emphasizing their effectiveness as sentinel species (Marneweck et al., 2021).
One key conservation strategy to slow down populations’ decline and mitigate human impacts, is the establishment and maintenance of protected areas (PAs), which can safeguard the most sensitive species. In this study, we investigated differences in the occupancy of mesocarnivore assemblages in two PAs under different management regimes – strict protection and sustainable use – in the northeast Atlantic Forest of Brazil. Combining camera-trapping and hierarchical community occupancy models, we estimated mesocarnivore species richness at the PA scale and assessed the influence of fine-scale environmental and anthropogenic factors on species-specific occupancy patterns. We empirically tested expectations that mesocarnivore occupancy is positively associated with i) strict levels of PA protection and larger size (Gray et al., 2016), ii) flat terrains (O’Malley et al., 2024) and iii) mature and better-preserved forest patches (Regolin et al., 2017), while being iv) negatively related to anthropogenic stressors (access roads and PAs limits; Goulart et al., 2009; Nagy-reis et al., 2017). However, species responses might be trait-dependent, including dependence on forest habitats, or being vulnerable or benefiting from human disturbance.
Materials and methodsStudy areasThe study was conducted in two PAs in southern Bahia, Brazil: Pau Brasil National Park (PNPB) and the Private Natural Heritage Reserve Estação Veracel (RPPNEV), both located in a biodiversity "hotpoint" within the Brazilian Atlantic Forest hotspot (Fig. 1). PNPB, a strictly PA, spans 19.027 ha, while RPPNEV, designated for sustainable use, covers 6.069 ha. These two PAs are key components of the Central Atlantic Forest Corridor, characterized by a super-humid climate with consistent rainfall year-round. The landscape features predominantly flat terrain, with elevations ranging from 10 to 100 m above sea level and includes mature vegetation, "muçununga" forests, seasonally or permanently flooded areas, and forests in various successional stages (CPRM, 2002). Poaching occurs in both PAs, while illegal logging mostly takes place within PNPB. Despite its lower legal protection status, RPPNEV benefits from a more intensive surveillance regime than PNPB, due to its smaller area and he greater number of personnel involved in active monitoring. The jaguar (Panthera onca), the apex predator of the Atlantic Forest, is virtually extinct in the region, while pumas (Puma concolor) persist in both areas (Magioli et al., 2023).
 and Private Reserve of Natural Heritage Estação Veracel (RPPNEV), southern Bahia, Brazil, depicting the location of the camera trap stations and main land uses (MapBiomas Project). The green area in the inset map represents the distribution of the Atlantic Forest biome.")
Location of the Pau Brasil National Park (PNPB) and Private Reserve of Natural Heritage Estação Veracel (RPPNEV), southern Bahia, Brazil, depicting the location of the camera trap stations and main land uses (MapBiomas Project). The green area in the inset map represents the distribution of the Atlantic Forest biome.
Mesocarnivores were surveyed using camera trapping, a common and versatile method for assessing the distribution of medium-sized mammals (Tobler et al., 2008). Sampling was conducted between 2018 and 2021, during four surveys lasting ∼60 days: three between August and October (2018, 2019, 2021) and one between April and June (2019). We used data collected at 34 sampling stations in PNPB and 27 in RPPNEV (Fig. 1). Stations were deployed with a spacing of 1.4 km, following the TEAM Protocol (Team, 2011), and located on dirt roads, trails, and inside the forest (i.e., outside roads and trails). At each station, a single camera trap (Bushnell Trophy Cam HD, Model 119876) was installed approximately 40 cm above the ground, and programmed to operate 24 h. Species-specific detection histories were generated using 3-day occasions. We organized mesocarnivore data in a stacked format, with each row representing the encounter history of an individual species at a specific site within one of the areas (PNPB or RPPNEV) and during a particular survey (1–4).
Community occupancy modelWe used a hierarchical community occupancy model (Dorazio and Royle, 2005) to assess changes in species-specific occupancy rates (i.e., the proportion of occupied/used area, proxy for local abundance; Steenweg et al., 2018) and heterogeneity in occupancy probabilities, while formally accounting for imperfect detection. The true occupancy of a species k at a site i, zki, is a Bernoulli random variable governed by occupancy probability ψki. We define occupancy as the probability of site use by a species during each survey period. Observations yki follow a Binomial distribution governed by detectability pki conditioned on the true occupancy state of each species (zki) and sampling effort (trap-days) at each site (Kki). We estimated species-specific occupancy and detection probabilities as random effects with area and survey-specific intercepts (β0,k,AreaSurvey[i]) to accommodate differences in baseline occupancy and detection probabilities across PAs, repeated surveys, and species. Despite losing the ability to distinguish the underlying temporal dynamics (i.e. colonization and extinction processes; MacKenzie et al., 2002), such formulation eases the estimation of model parameters when analyzing sparse datasets. For inferences on species richness (Wki), we augmented the observation data with all-zero observations (Dorazio and Royle, 2005) of hypothetical species up to a total of 11 mesocarnivores that may occur in the study region following the complete list of mammals in the study areas (Magioli et al., 2023). We estimated a latent binary indicator variable wi,Area indicating whether or not each species is part of the community in each PA.
Under this base formulation, we implemented two separate models. First, we estimated species richness and species-specific occupancy (Ψ) and detection probabilities (p) using only area-by-survey intercepts. We opted not to include covariates in this initial model due to the limited data available for multiple species (< 5 independent records, one-hour interval), hindering parameter estimation. The model can be described as:
Second, we considered only species with detection values higher than 0.1 in all surveys at both areas (see Results) and modeled occupancy and detection parameters as a logit-linear function of environmental and anthropogenic disturbance covariates. We modeled species-specific site occupancy probability as a function of: (i) tree basal area (BASAL), measured in the field as the sum of trees’ basal area with a circumference greater than 15 cm in a 50 × 3 m cross plots around each camera-site (Appendix 1); (ii) terrain slope at the camera-site (SLOPE), extracted from a 30 m resolution digital elevation model (Shuttle Radar Topography Mission, https://earthexplorer.usgs.gov/), and; (iii) distances to the nearest PA limits (LIMIT) and (iv) dirt roads (ROAD), measured in QGIS 3.28.3 from remote-sensed imagery in Google Earth (Appendix 2). For detection probability, we considered the effect of fine-scale camera placement (CAMPLACE) using a categorical covariate distinguishing sites placed at roads or wildlife trails from sites placed inside the forest matrix as the reference category.. Prior to the analysis, we ensured that there was no significant collinearity among covariates (|r| > 0.7) and standardized them to have a mean of 0 and a standard deviation of 1. We modeled species-specific parameters as random effects from a common underlying distribution governed by community-wide hyperparameters. We followed Zipkin et al. (2009) and fit a single global model with a limited number of covariates for which there was a strong a priori justification; such that:
We analyzed the models using Markov Chain Monte Carlo (MCMC) simulations in JAGS, implemented through jagsUI package (version 1.5.2) in software R (version 2023.06.1 + 524 R Core Team). We used three chains of 50,000 iterations, with a burn-in of 10,000, and retaining every 10th iteration. We assessed chain convergence based on a Gelman–Rubin statistic < 1.1 (Gelman et al., 2021) and visual inspection of trace plots. Model fit was evaluated by estimating the discrepancy between the deviance residuals of the observed and simulated data from the fitted model (Broms et al., 2016), suggesting reasonable fit (Bayesian p-values of 0.495 and 0.503 for each stage, respectively).
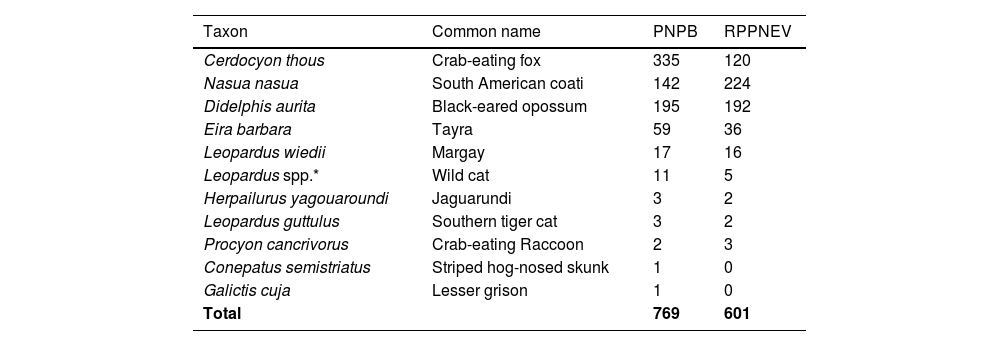
ResultsWe obtained 1310 independent records of 11 mesocarnivore species over a total of 13,609 trap-days (Table 1, Appendix 3), all expected to occur in the study region, except Leopardus pardalis (Magioli et al., 2023). Mesocarnivore species richness was higher in PNPB (11) than in RPPNEV (9), as estimated from the first-stage model with all species included. Two mesocarnivore species were exclusively detected in PNPB: Conepatus semistriatus and Galictis cuja, albeit with only one detection each.
Mesocarnivore species recorded in Pau Brasil National Park (PNPB) and Private Reserve of Natural Heritage Estação Veracel (RPPNEV), southern Bahia, Brazil, depicting the number of independent records of each species per area. Leopardus spp.* corresponds to unidentified records of L. guttulus or L. wiedii.
| Taxon | Common name | PNPB | RPPNEV |
|---|---|---|---|
| Cerdocyon thous | Crab-eating fox | 335 | 120 |
| Nasua nasua | South American coati | 142 | 224 |
| Didelphis aurita | Black-eared opossum | 195 | 192 |
| Eira barbara | Tayra | 59 | 36 |
| Leopardus wiedii | Margay | 17 | 16 |
| Leopardus spp.* | Wild cat | 11 | 5 |
| Herpailurus yagouaroundi | Jaguarundi | 3 | 2 |
| Leopardus guttulus | Southern tiger cat | 3 | 2 |
| Procyon cancrivorus | Crab-eating Raccoon | 2 | 3 |
| Conepatus semistriatus | Striped hog-nosed skunk | 1 | 0 |
| Galictis cuja | Lesser grison | 1 | 0 |
| Total | 769 | 601 |
Detection probability estimates at the community level was 0.045 (Global mean across all species, areas, and surveys), although there was considerable interspecific and temporal variation (Appendix 4a). Five mesocarnivore species - C. semistriatus, G. cuja, Procyon cancrivorus, Herpailurus yagouaroundi, and Leopardus guttulus – were detected few times (<5) in each area, resulting in detection probabilities below 0.1 throughout all surveys and unprecise occupancy estimates (Appendix 4b). As such, four species were considered in the second-stage model for inferences on mesocarnivore distribution – Cerdocyon thous, Didelphis aurita, Eira barbara, and Nasua nasua – and data for all Leopardus species were combined at the genus level, including detections with uncertain phenotypic differentiation, and modeled as a distinct functional species.
The occupancy probability of the species did not show obvious differences between PAs (Fig. 2), and we observed a high degree of interspecific and temporal variation across surveys, which was often asymmetric between areas. Nasua nasua was more widespread in RPPNEV during all surveys (Ψ > 0.75). Didelphis aurita had higher occupancy in PNPB in surveys 2 and 3 (Ψ = 0.788 and 0.750, respectively), E. barbara showed an increased occupancy in PNPB during surveys 3 and 4 (Ψ = 0.573 and 0.679, respectively), and Leopardus sp. had higher occupancy in PNPB in surveys 2 and 4 (Ψ = 0.661 and 0.431, respectively) and in RPPNEV during survey 3 (Ψ = 0.612).
, survey 2 (April–June/2019), survey 3 (August–October/2019), and survey 4 (August–October/2021), conducted in Pau Brasil National Park (PNPB) and Private Reserve of Natural Heritage Estação Veracel (RPPNEV), southern Bahia, Brazil. Points above the diagonal line indicate greater species occupancy in the RPPNEV (y-axis), while those below it indicate higher species occupancy in the PNPB (x-axis). The error bars depict 95% Bayesian credible intervals. Species codes: crdt- Cerdocyon thous; ddla-Didelphis aurita; erbr- Eira Barbara; lsp-Leopardus sp.; nsns-Nasua nasua.")
Species-specific occupancy probability estimated in the second stage of the model in each survey: survey 1 (August–October/2018), survey 2 (April–June/2019), survey 3 (August–October/2019), and survey 4 (August–October/2021), conducted in Pau Brasil National Park (PNPB) and Private Reserve of Natural Heritage Estação Veracel (RPPNEV), southern Bahia, Brazil. Points above the diagonal line indicate greater species occupancy in the RPPNEV (y-axis), while those below it indicate higher species occupancy in the PNPB (x-axis). The error bars depict 95% Bayesian credible intervals. Species codes: crdt- Cerdocyon thous; ddla-Didelphis aurita; erbr- Eira Barbara; lsp-Leopardus sp.; nsns-Nasua nasua.
No clear occupancy response to site-scale covariates was observed at the community level, with mesocarnivore species often exhibiting contrasting associations with environmental and disturbance factors. Cerdocyon thous occupancy was strongly associated with more disturbed forest sites, i.e. with a lower basal area (βBASAL = −0.97, [range: −1.85 to −0.19]; Fig. 3A), closer to roads (βROAD = −0.57 [−1.33 to 0.05]; Fig. 3C). Conversely, N. nasua occupancy moderately increased in sites with higher basal area (βBASAL = 0.42 [−0.12 to 1.04]; Fig. 3A) and that of D. aurita moderately increased further away from roads (βROAD = 0.41 [−0.04 to 0.90]; Fig. 3C). The occupancy of N. nasua and Leopardus sp. was positively associated withthe proximity to PA limits (βLIMITS = −0.51, [−1.45 to 0.05] and βLIMITS = −0.28 [−0.70 to 0.09], respectively; Fig. 3D). None of the species had their occupancy substantially influenced by terrain slope (Fig. 3B), and E. barbara did not respond to any covariate. Detection probability for most mesocarnivores was strongly associated with camera-trap placement (Fig. 3E), with bimodal responses across species. While detectability of N. nasua and E. barbara was higher in the forest (βCAMPLACE = −0.38 [−0.68 to −0.08] and βCAMPLACE = −0.98 [−1.77 to −0.19], respectively), the detection of C. thous and Leopardus sp. increased in roads or trails [βCAMPLACE = 3.78 [2.89–4.72] and β CAMPLACE = 0.19 [−0.13 to 0.49], respectively).
 and detectability (E) probabilities for the five selected species in Pau Brasil National Park (PNPB) and Private Reserve of Natural Heritage Estação Veracel (RPPNEV), southern Bahia, Brazil. Points are posterior medians and error bars represent 95% Bayesian credible intervals (BCI). Colors indicate the direction of the effect (negative in red, positive in green, no effect in gray), while the shade of red and green colors reflect the probability of an effect, calculated as the proportion of the posterior with the same sign as the mean: darker tones indicate strong support (≥0.95) and lighter tones indicate moderate support (≥0.90). Vertical lines depict mean (solid line) and 95% BCI (dashed lines) community effects.")
Effect size of site-scale environmental and disturbance covariates on occupancy (A-D) and detectability (E) probabilities for the five selected species in Pau Brasil National Park (PNPB) and Private Reserve of Natural Heritage Estação Veracel (RPPNEV), southern Bahia, Brazil. Points are posterior medians and error bars represent 95% Bayesian credible intervals (BCI). Colors indicate the direction of the effect (negative in red, positive in green, no effect in gray), while the shade of red and green colors reflect the probability of an effect, calculated as the proportion of the posterior with the same sign as the mean: darker tones indicate strong support (≥0.95) and lighter tones indicate moderate support (≥0.90). Vertical lines depict mean (solid line) and 95% BCI (dashed lines) community effects.
Our findings revealed that the mesocarnivore community structures in PNPB and RPPNEV are complex and heterogeneous. Both PAs support a diverse range of species, with PNPB exhibiting greater species richness. Overall, species detectability was low, and occupancy showed strong interspecific and temporal variation. There were no uniform response to the environmental and anthropogenic factors at the assemblage level l; with contrasting patterns emphasizing the importance of species-specific evaluations. In general, environmental factors induced stronger but fewer responses in site occupancy by mesocarnivores, while anthropogenic factors seemed to cause greater variation in habitat use, potentially facilitating species occupancy. Notably, there was a tendency for species to occupy the borders of the PAs, likely benefiting from foraging opportunities in adjacent agricultural areas.
Despite the relatively lower abundance of carnivores compared to other trophic groups (Newbold et al., 2020), the low detectability observed in our study compared to other areas in the Brazilian Atlantic Forest, such as Iguacu National Park (da Silva et al., 2018) and Vale Natural Reserve (Wolff et al., 2019), suggests low population abundances within the studied areas (Burns et al., 2019). The detectability variation in response to camera trap location was consistent with other studies in the Atlantic Forest (Di Bitetti et al., 2010). The higher species richness in PNPB is likely due to its larger size—which supports more habitats, greater resource availability, and larger populations (Connor and McCoy, 2001)—rather than its legal protection status, as the sustainable use PA has an intensive surveillance system, a pattern already observed in southern Bahia (Magioli et al., 2021b). Furthermore, the higher species occupancy in PNPB underscores the critical role of protecting large areas for mesocarnivore conservation in the Atlantic Forest.
The temporal variation in species-specific occupancy across both PAs is particularly intriguing for a relatively stable tropical forest. Such differences can be partly explained by low precision of occupancy estimations (a season's credible interval overlaps the mean of other seasons), but may also reflect natural population fluctuations combined with human interference. Notably, the most variable occupancy estimation was observed for D. aurita, the smallest species among those investigated, whose life cycles are shorter and reproductive rates are higher (Emmons and Feer, 1997), traits often associated with faster population dynamics.
Influence of environmental and anthropogenic factorsNatural factors generated stronger but fewer responses in site occupancy by mesocarnivores, which may result from the relatively low environmental variability in the study areas. Despite harboring forests with distinct disturbance histories, the PAs have relatively flat terrain and low edaphic variation. The basal area, a locally measured variable representing mature forest, influenced two mesocarnivore species, indicating that it may be a reliable predictor for this group and other species, and should be incorporated in future studies. The increase in basal area was important for N. nasua habitat use, which aligns with their preference for forested environments (e.g., Dutra et al., 2023). Conversely, C. thous appears to benefit from the reduction in basal area, which is expected since this habitat and diet generalist species thrives in more open and disturbed environments (Lucherini, 2015). No species exhibited a response to terrain slope, which is potentially attributed to the intricate interactions between terrain and resource availability. While flatter terrains tend to have higher prey abundance (de Oliveira et al., 2010), steeper terrains are strongly associated with water drainage in the study areas, a widespread resource across both PAs. Moreover, slope may play a more influential role for carnivore occupancy in areas with more rugged topography, such as mountainous regions (Nisi et al., 2022).
Anthropogenic factors influenced the habitat use of a greater number of mesocarnivore species. The higher occupancy of C. thous near dirt roads was expected given its tolerance to disturbed and open areas, likely benefiting from the surrounding agricultural areas to forage (Monteiro-Alves et al., 2018). The opposite response of D. aurita can be related to the fact that the species depend on forested habitats due to its scansorial habit (Emmons and Feer, 1997), with roads potentially increasing its exposure to predators, for example. Contrary to our initial expectation, the increasing distance to PA limits decreased the occupancy of Leopardus sp. and N. nasua. While sites closer to the PA limits face more disturbances, adjacent lands dominated by eucalyptus, pasture, and agriculture can provide direct resources for mesocarnivores and serve as alternative habitats for small vertebrates (e.g., rodents), which are prey for these species, creating foraging opportunities near forest edges (Di Bitetti et al., 2010; Massara et al., 2018).
The heterogeneity in responses to the covariates was anticipated, as mesocarnivores often exhibit distinct habitat specializations. It is evident that some species, like C. thous, benefit from disturbed areas, while others, such as E. barbara, are negatively affected by these conditions. In the context of increasing anthropogenic disturbances, this may lead to a "winners/losers" dynamic (Baskin, 1998) within the study area. A key consequence of this shift is the alteration of the trophic chain and the disruption of ecological functions (Roemer et al., 2009). It is important to note that mesocarnivores play diverse ecological roles; for example, C. thous and D. aurita are important seed dispersers, while Leopardus sp. exert a top-down control of rodent populations (Magioli et al., 2021a).
The few strong responses to environmental and anthropogenic factors, and the low overall occupancy rates in the study areas, indicate that other factors may be influencing the mesocarnivore assemblage, such as food availability and/or intra-guild interactions (Di Bitetti et al., 2010). The absence of L. pardalis, the largest mesopredator in this region, was unexpected—although the species has been poorly recorded in previous surveys across the Atlantic Forest of southern Bahia (Magioli et al., 2021b). The lack of this species may produce the 'pardalis effect', in which the absence of L. pardalis leads to increased abundance of smaller felids such as L. wiedii and L. guttulus (Silva-Magana and Santos-Moreno, 2020). The fragmentation of the Atlantic Forest into small, isolated patches, coupled with the poorly connection of PAs network, particularly in northeastern Brazil, also likely contributes to the generally low occupancy rates observed.
ConclusionsOur study highlights the complex and heterogeneous mesocarnivore assemblage supported by the studied PAs, but also stresses the effect of anthropogenic factors on species occupancy. The low detectability and occupancy of the mesocarnivores in our study compared to other areas in the Atlantic Forest implies either naturally low abundance within these PAs, or other limiting factors impacting these populations. The absence of assemblage-wide responses to environmental and anthropogenic factors underscores the importance of species-specific strategies for species conservation. Interestingly, some species tended to use the edges of the PAs, suggesting the use of agricultural areas as supplementary food resources and part of their habitat (Magioli et al., 2019). These findings, coupled with the absences of the apex predator (P. onca) and the largest mesopredator (L. pardalis), along with and the presence of species typically associated with open areas (C. semistriatus), suggest that anthropogenic disturbances in both areas have shaped the structure of local mesocarnivore assemblages.
CRediT authorship contribution statementMarília Marques: Writing - original draft, Visualization, Formal analysis, Data curation. Marcelo Magioli: Writing - review & editing, Supervision, Data curation. Pedro Monterroso: Writing - review & editing, Supervision, Formal analysis. Gonçalo Curveira-Santos: Writing - review & editing, Formal analysis. Camila Righetto Cassano: Writing - review & editing, Supervision.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We acknowledge the financial support from the Pró-Reitoria de Pesquisa e Pós-Graduação (PROPP/UESC:073.6764.2020.0003917-18) and Coordination for the Improvement of Higher Education Personnel (CAPES); M. Marques scholarship was financed by CAPES. M. Magioli thanks the São Paulo Research Foundation (FAPESP) for the post-doctoral scholarship (#2022/06791-9|). G.Curveira-Santos. was funded by Fundacão para a Ciência e a Tecnologia (FCT) in the frame of Individual Call to Scientific Employment Stimulus program [2023.08961.CEECIND/CP2845/CT0007].
The following is Supplementary data to this article:

