

In this article we investigate how invasion by Pinus taeda L. alters plant community composition and functional traits in sand dunes. In invaded psammophilous vegetation in southern Brazil, we randomly selected 15 P. taeda individuals and measured their age based on the number of growth rings. For each tree, we sampled 1m2 quadrats beneath and 2m away of its crown. In each quadrat, we assessed non-graminoid plant composition and performed a multivariate analysis of variance. Data on species’ functional traits were scaled up to community level, and their variations were compared to tree age by linear regression. Species’ composition and functional traits under P. taeda crown were different from control plots. Tree age affected the presence of thorns, potential height, and functional diversity. P. taeda individuals likely change the micro-habitat beneath their crowns, thereby benefiting species with some traits and increasing functional diversity of plants that are not characteristic of open dune communities.
© 2014 Associação Brasileira de Ciência Ecológica e Conservação. Published by Elsevier Editora Ltda.
It is well documented that Pinus species are invasive worldwide (Richardson 2006). It has been found that some life history traits, such as short juvenile period and numerous small winged seeds, which characterize these species as pioneers in their native range, are also responsible for their invasiveness (Richardson 2006; Rejmánek 1996). Furthermore, Pinus invasion is more prevalent in low biomass ecosystems and communities with less competition with other tree species (Richardson & Higgins 1998). These characteristics make dune ecosystems, grasslands, and shrublands the most vulnerable environments to invasion (Richardson & Higgins 1998).
Dune ecosystems are characterized by high insolation and strong winds, strongly influenced by the sea and by severe disturbances, such as droughts and sand burial (Maun 2009). Dune species usually present characteristics of early successional stages, such as rapid colonization and great dispersal ability, while not adapted to low light conditions and competition (Tilman 1985). Therefore, dune communities are more susceptible to invasion by better competitors that are able to cope with such limiting abiotic conditions.
It is known that Pinus invasion can influence ecosystem processes by increasing shading and litter amount (Craine & Orians 2004). The release of leaf allelochemicals or root exudates may inhibit the development of plants under the crown (Fernandez et al. 2006). On its natural range, Pinus species are important in the ecological succession, from open dunes to woodlands, by means of nucleation (Maun 2009). So it is expected that a similar process may occur at the same kind of environment where Pinus is invasive, representing a new functional group introduced in the community and thus changing important ecosystem functions.
Changes in ecosystem processes can also arise from changes in the plant community and its functional patterns (Violle et al. 2007). Functional patterns are usually given by the species’ functional traits. This approach is used for answering questions in community ecology that cannot be well treated using ordinary parameters such as species diversity and composition.
In the present article, we analyze the effect of Pinus taeda invasion on plant communities of coastal sand dunes. First, by determining the age P. taeda individuals, we were able to analyze the community shifts actually caused by the invader and not due to preexisting environmental conditions that affect both native and exotic species. By comparing communities under P. taeda trees of different ages, we investigated how time span since tree establishment is related to species composition shifts. Afterwards, a formal methodology was adopted for analyzing changes on the functional structure of plant community under invasion, evaluating convergence and divergence assembly patterns.
Material and methodsThe study was performed at the Itapeva State Park (29°22’22”S, 49°45’06”W) in the highly endangered and vulnerable coastal plain of southern Brazil (SEMA 2006). The selected area had psammophilous vegetation on lowlands between the fore dunes and inland mobile dunes. P. taeda occurrence in the park is cited by SEMA (2006), but no precise information is given on how it was introduced. However, a population pattern of sparse individuals of many different ages indicates that the introduction was caused by invasion from adjacent areas.
We established a 100×100m grid and, at each grid node (from a total of 131), we measured the tree height of all P. taeda individuals within a 25-meter radius from the node. The mapped P. taeda were stratified into three height classes (lower than 1m, between 1 and 3m, and more than 3m), and five individuals were randomly sampled per stratum, totalizing 15 individuals. We determined the age of all 15 sampled P. taeda by trunk sampling and tree ring counting (Stokes & Smiley 1968).
We marked a 1×1m plot under the crown of each sampled P. taeda and paired control plots of the same size, randomly located 2m away from the crown limit of each tree (15 pairs). Preliminary observations indicated that the P. taeda trees in the dunes were less vigorous and had more open canopies than those in more benign environments, and there was no evidence of needle accumulation at 2m away from the trees. Thus, we considered this distance sufficient to distinguish invaded from non-invaded communities, and closer plots more likely had similar environmental conditions before the invasion than distant plots.
The presence of all non-graminoid species in each plot was recorded. We used presence/absence data in accordance with Wilson (2012), who demonstrated that species incidence may give equal or better results than abundance data.
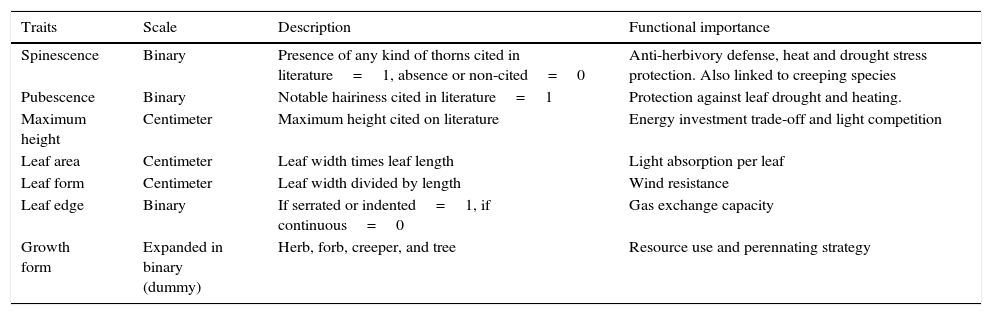
Data on species’ functional traits were compiled from the scientific literature, information from specialists, and observation of herbarium specimens. The selected traits (linked to survival strategies) and their functional importance are shown in Table 1.
Traits used for the description of species found at psammophilous vegetation under P. taeda individuals. Functional importance follows Pérez-Harguindeguy et al. (2013).
| Traits | Scale | Description | Functional importance |
|---|---|---|---|
| Spinescence | Binary | Presence of any kind of thorns cited in literature=1, absence or non-cited=0 | Anti-herbivory defense, heat and drought stress protection. Also linked to creeping species |
| Pubescence | Binary | Notable hairiness cited in literature=1 | Protection against leaf drought and heating. |
| Maximum height | Centimeter | Maximum height cited on literature | Energy investment trade-off and light competition |
| Leaf area | Centimeter | Leaf width times leaf length | Light absorption per leaf |
| Leaf form | Centimeter | Leaf width divided by length | Wind resistance |
| Leaf edge | Binary | If serrated or indented=1, if continuous=0 | Gas exchange capacity |
| Growth form | Expanded in binary (dummy) | Herb, forb, creeper, and tree | Resource use and perennating strategy |
For the evaluation of an overall effect of P. taeda in community composition and functional traits, we performed a multivariate analysis of variance (MANOVA) with permutation test (Pillar & Orlóci 1996, Anderson 2001) comparing P. taeda to control plots. Euclidean distances, restricting permutations within paired plots (Pillar & Orlóci 1996), were used for this purpose. Compositional differences were accessed directly on presence/absence data matrix, while functional differences were based on species traits scaled up to the community level (Pillar et al. 2009).
For accessing whether possible compositional differences between plots under P. taeda and the controls were due to P. taeda influence or to its preferential colonization in certain communities, we tested whether the compositional Euclidean distance between P. taeda and the paired control plot was explained by tree age using linear models with permutation tests (Manly 1991). Both analyses were performed with the software MULTIV (Pillar 1997).
We also examined functional community patterns related to P. taeda age taken as a proxy for community development. The analysis was based in traits of species occurring in P. taeda plots only (control plots were not considered). We performed the procedure described in Pillar et al. (2009) (more details in Appendix I) to identify traits better representing variation in response to the tree age gradient in terms of convergence and divergence patterns. For the selected convergence traits, we used simple regressions on tree age of the community-weighted means, separately by trait. For the selected divergence traits, we used regression to access how functional diversity (Rao’s Quadratic Entropy, Rao 1982) varied, with tree age as a predictor.
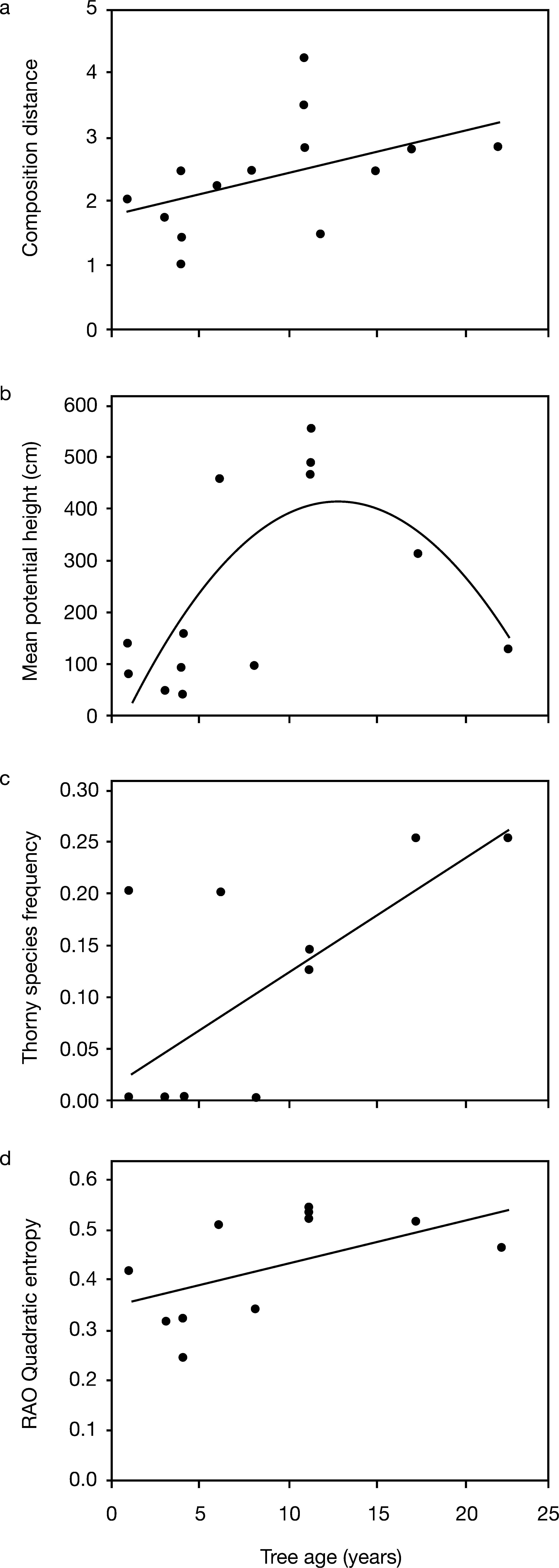
ResultsThe age of the P. taeda trees varied between 1 and 20 years and height varied between 1.3 and 4.5m. A total of 47 non-graminoids species was recorded among the plots (see Appendix II for the floristic list). Two of the plots under P. taeda trees lacked vegetation. Some species were common in plots under P. taeda while infrequent in control plots. Other species were more common only in control plots (Fig. 1). It is important to highlight that all tree species found on the sampled plots were seedlings or saplings. We found a significant difference in species composition between plots under P. taeda and control plots (P=0.005, MANOVA using permutation test). This difference increased with age of the P. taeda tree, as shown by the linear regression (R2=0.23, P=0.065; Fig. 2a).
 and control plots (n=15). Only species with frequency higher than 20% in any treatment are shown.")
 composition Euclidean distance between paired P. taeda and control plots (R2=0.23, P=0.065); (b) community mean for species potential height (R2=0.56, P=0.02); (c) community mean for presence of thorns (R2=0.450, P=0.01); (d) Rao quadratic entropy (for the traits shrub and tree growth forms, pubescence, potential height, leaf form, and presence of thorns) (R2=0.308, P=0.047).")
Relationships assessed by linear regressions between P. taeda tree age and community characteristics: (a) composition Euclidean distance between paired P. taeda and control plots (R2=0.23, P=0.065); (b) community mean for species potential height (R2=0.56, P=0.02); (c) community mean for presence of thorns (R2=0.450, P=0.01); (d) Rao quadratic entropy (for the traits shrub and tree growth forms, pubescence, potential height, leaf form, and presence of thorns) (R2=0.308, P=0.047).
Plots described by community weighted mean traits also showed a significant difference (using MANOVA) in trait values between plots under P. taeda and control plots (P=0.052). Potential height and presence of thorns were traits that jointly expressed significant trait-convergence related to tree age (ρ(TE)=0.43, P=0.047). Regression models of these traits on tree age (Figs. 2b and 2c) demonstrated that the presence of thorns was linearly and positively related to the tree age gradient, while species potential height had the highest values on intermediate ages.
Shrub and tree growth forms, pubescence, potential height, leaf form, and presence of thorns jointly expressed significant trait-divergence patterns related to tree age. Functional diversity was positively related to tree age, as indicated by the regression of Rao quadratic entropy (calculated only for these traits) on tree age (Fig. 2d).
DiscussionOur results indicate that P. taeda trees indeed affect the species composition in the psammophilous plant communities under their crown (Figs. 1 and 2a). Moreover, the effect is also noticeable in the species’ functional traits. P. taeda, as a unique invading tree in this dune habitat, produces changes in the ecosystem. These changes are caused not only by the new growth form added, but also by the effects of the tree on the native community under its crown.
It can be stated that P. taeda trees caused the perceived differences, since the changes were only noticeable directly below the crown of P. taeda and not in control plots 2m apart from the tree crowns. Moreover, the differences between species composition under the tree crowns and the adjacent plot increased with increasing tree age (Fig. 2a). Thus, tree age is a significant factor in altering community composition. While plots under young trees presented almost no difference from control plots, the plots under old trees differed greatly from the control, as observed in the linear regression (Fig. 2a).
Although we did not test for causes of this differentiation, P. taeda likely generates a microenvironment that protects understory plants from high intensities of sun radiation and wind (Craine & Orians 2004), acting as a nucleation agent (Maun 2009). The older the tree, the more shade and wind shelter it provides, therefore favoring shade-tolerant species (such as Rumohra adiantiformis) and creeping plants that require a more advanced woody environment (such as Smilax campestris) (Fig. 1). The presence of P. taeda also appears to inhibit the growth of species that depend on more intense sunlight, such as Tibouchina urbanii and Asteraceae species (e.g., Pterocaulon angustifolium and Baccharis radicans; Fig. 1). Another important factor that may cause changes of plant communities under Pinus spp. is the production and accumulation of needles. Pinus spp. litter is difficult to decompose and causes the accumulation of a thick layer, which modifies the soil of the dunes, retaining more nutrients and soil moisture at the local scale (Craine & Orians 2004). Furthermore, not only the presence of the tree itself, but also the time extent of its presence as a cumulative effect, may explain the effects of the tree on the community under the crown.
Tree age also affected the functional traits composition in the communities under the crown, favoring plants with spines (Fig. 2c), while potentially taller plants (greater maximum height) are favored under trees of intermediate age (Fig. 2b). Tree age also increased functional diversity when considering shrub and tree growth forms, pubescence, potential height, leaf form, and presence of thorns (Fig. 2d). These results demonstrate that changes observed in plant composition can be also perceived in species’ functional traits. Thus, it can be inferred that changes caused by P. taeda individuals are selecting the replacement of dune species by species with different attributes that are more compatible for survival in the new environment.
One important trait is the presence of thorns, often associated with creeping species, such as Smilax campestris, common at plots under P. taeda crowns (Fig. 1). Another relevant trait related to community development, indicated by P. taeda age, was the potential height of the species established under the crown. P. taeda trees may provide perches for birds, allowing for seed dispersal of tree species and their establishment (such as Myrsine parvifolia and Sapium glandulosum, both with fleshy fruits dispersed by animals). However, after a certain age (approximately 13 years), P. taeda appears to negatively affect tall species, perhaps due to the cumulative effects of needle and exudates that exclude some tree species. In terms of conservation or management, this process should not be perceived as positive for psammophilous environments: the species established under the crowns are not the same species of the psammophilous initial vegetation. Moreover, many P. taeda individuals can die before reverting the effects under tall species, so it will act as a nucleator plant, changing psammophilous open vegetation into a woody one.
The observed increase of functional diversity may be indicating that under P. taeda crown other plant functional types are able to survive. Abiotic filters are stressful at open sand dunes (Maun 2009) and P. taeda trees may be generating new non-stressful microhabitats, protected from too much wind and sun radiation, hence allowing the establishment of species (from other vegetation types of the park) with a wider range of functional traits in the community. Therefore, though the functional diversity in the psammophylous area may increase with P. taeda invasion, the overall diversity may decrease due to the loss of psammoplhylous open vegetation inside the park.
This study demonstrated that P. taeda can have significant impacts on psammophilous plant community composition and functional traits, when considering non-graminoid species. As the invasion proceeds altering these natural areas, open vegetation species will be restricted to smaller areas, changing not only species composition but also the vegetation type itself. As many other coastal ecosystems, psammophilous vegetation is endangered in Southern Brazil, and the genus Pinus is highly invasive in these ecosystems (Bechara et al 2013). Managers and policymakers must be aware of the risks of an unnatural shift to woody vegetation in this type of ecosystem.

