

Free-roaming domestic cats can negatively affect wildlife, and understanding how they use space is fundamental to predict their potential impacts. In this study we describe the use of space of 48 owned free-roaming cats and assess the level of care provided by owners in rural areas near priority conservation areas in southern Chile. Additionally, we use camera trap data from two protected areas to assess spatial overlap with wild vertebrates, particularly the guigna (Leopardus guigna), a vulnerable forest-dwelling felid. Cat home ranges were variable (1–47.2 hectares) and their activity was mostly concentrated within 100 meters of the household, with maximum foray distances up to 2.5km. Cats used wooded areas (12% of fixes on average), mostly native forest, where spatial overlap with endemic and/or threatened species was documented. Nonetheless cats were infrequently detected in nearby protected areas (1.7% of GPS locations and 1% of the cameras). The use of wooded areas was strongly associated with the distance from the household to the forest edge and home range areas were negatively associated with house density. Our findings suggest that in areas isolated from households (>200m), the problem of cats is marginal and restricted to the edges. Management was in general deficient, observing birth control in only 13% of cats and lack of preventive health care in 86% of cats. Cat impacts could be exacerbated under increasing land subdivision in southern Chile where, without population control strategies, cats may become an emergent threat for native wildlife.
The domestic cat is one of the most abundant carnivores in the world (Serpell, 2019), and one of the most popular pets. Listed as one of the 100 worst invasive species in the world (Lowe et al., 2000), the domestic cat can exert different negative effects on wildlife, including predation, competition, hybridization and pathogen transmission (Medina et al., 2011; Loss et al., 2013; Medina et al., 2014, Doherty et al., 2016). Cat impacts are especially prominent on islands, where they are a major driver of vertebrate extinction (Medina et al., 2011, Medina et al., 2014). Although feral and stray cats are responsible for an important proportion of the impacts on biodiversity (Medina et al., 2011; Loss et al., 2013), owned cats are also of concern, as they are often allowed to range free (Kays and DeWan, 2004; Ferreira et al., 2011; Loss et al., 2013; Loss and Marra, 2017).
The assessment of the spatial use of free-roaming cats is useful to locate and estimate the extension of their area of influence (Van Heezik et al., 2010; Hervías et al., 2014), to identify for example, the spatial extent over which prey are encountered and exposed to cat predation (Hervías et al., 2014). Furthermore, given that domestic cats may represent a risk for native carnivores through disease spillover (Chalkowski et al., 2019, Filoni et al., 2011; Millán and Blasco-Costa, 2012; Mora et al., 2015; Veronesi et al., 2016), understanding their use of space is important to identify factors that may facilitate contact with native hosts and elucidate pathogen transmission risk and patterns (e.g. Woodroffe and Donnelly, 2011; Sepúlveda et al., 2014).
Habitat use by domestic cats has been described in numerous studies. Home ranges are highly variable, ranging from <1 to 300ha (Barrat, 1997; Metsers et al., 2010; Van Heezik et al., 2010; Gehrt et al., 2013). Home range has been associated with the availability and distribution of food resources, predation risk and ownership (Horn et al., 2011; Gehrt et al., 2013). For example, unowned cats have been reported to have larger home ranges than owned cats (Horn et al., 2011). Home range size also differs by sex; male home ranges are described to be larger (Wierzbowska et al., 2012; Gehrt et al., 2013). It has also been reported that domestic cats prefer areas near human settlements and roads, whereas remote areas or forests are used less frequently (Ferreira et al., 2011). A recent meta-analysis described sex, age and house density as the main factors influencing home range of domestic cats (Hall et al., 2016).
Few studies in Chile have provided insights into the ecology of domestic cats (but see Silva-Rodríguez and Sieving, 2011, Schüttler et al., 2018, Escobar-Aguirre et al., 2019, Silva-Rodríguez et al., 2020). Cats in rural areas are commonly owned not only as pets, but also to control rodents (Silva-Rodríguez and Sieving, 2011, Schüttler et al., 2018). Supporting this role, Muñoz-Zanzi et al. (2014) found that rodent counts and the number of cats were negatively correlated. Free-roaming domestic cats are common in human settlements located adjacent to natural areas inhabited by endemic and/or threatened vertebrates, representing a potential threat (Silva-Rodríguez et al., 2019). A previous study showed that 89% of cat owners reported their cats preyed on native wildlife, including endemic small mammals and native birds (Silva-Rodríguez and Sieving, 2011). Pathogens linked to domestic cats such as feline leukemia virus (FeLV) and feline immunodeficiency virus (FIV) have been described infecting populations of the threatened wild cat guigna (Leopardus guigna) in central and southern Chile (Mora et al., 2015, Sacristán et al., 2021) and co-occurrence of these felids has been reported in urban and peri-urban areas of southern Chile (Silva-Rodríguez et al., 2020). Potential impacts of cats on native wildlife could be favored by the fact that cats are often poorly managed (Silva-Rodríguez and Sieving, 2011); despite these concerns, very little is known regarding their use of space in priority conservation areas in Chile.
Our aim was to investigate the use of space by rural free-roaming domestic cats, focusing on the conservation implications for vertebrates that inhabit the Valdivian rainforest ecoregion, recognized as a biodiversity hotspot (Myers et al., 2000). Potential spatial overlap with the guigna, a vulnerable forest-dwelling felid, was assessed, since disease transmission by domestic cats has been described as an emerging threat for guignas (Busch et al., 2021; Mora et al., 2015; Sacristán et al., 2019, 2021; Sieg et al., 2020; Napolitano et al., 2015). We characterized local domestic cat populations in the study sites in terms of demography and care provided by owners, to identify potential risk factors or threats to wildlife. We hypothesized that domestic cats roamed away from their households using native forest, facilitating encounters and interactions with native fauna. We discuss some of the possible mitigation measures to reduce the impact of rural domestic cats near conservation areas in Chile. To our knowledge, this is the first study assessing the use of space of free-roaming domestic cat populations and their potential spatial overlap with native and threatened wildlife in Chile.
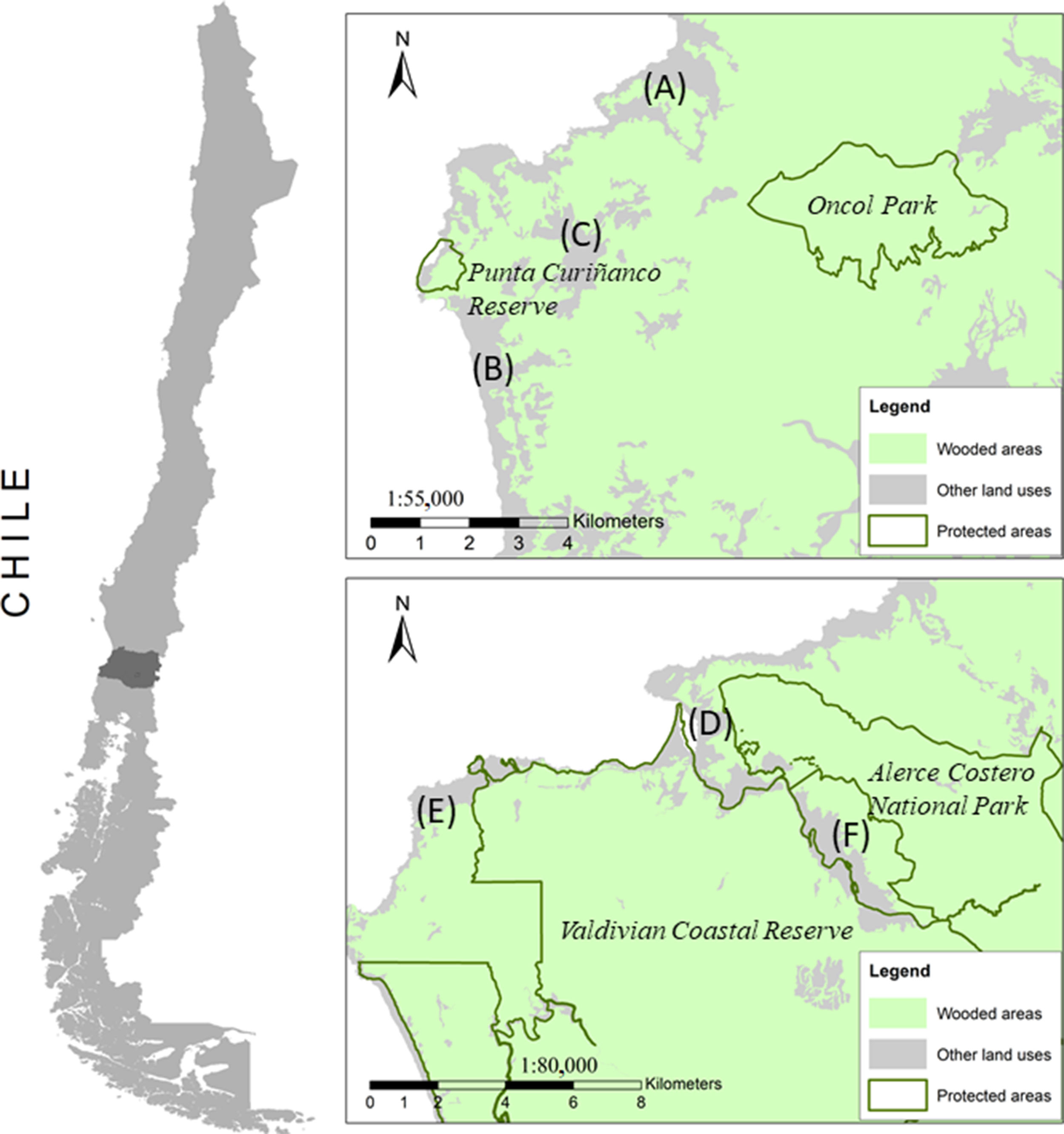
MethodsStudy areaThe study was conducted in the Coastal Range of Los Ríos region in southern Chile, an ecoregion characterized by high levels of endemism (Armesto et al., 1998) and considered as a global priority area for biodiversity conservation (Myers et al., 2000, Olson and Dinerstein, 2002). The study area included six rural communities (Chaihuín, Huiro, Cadillal, Curiñanco, Pilolcura and Bonifacio) adjacent (0.2–7.0km) to four protected areas including Alerce Costero National Park (24,649ha, administered by CONAF, the Chilean Government Protected Areas Administration office), the Valdivian Coastal Reserve (50,808ha, administered by The Nature Conservancy), Oncol Park (754ha, administered by Arauco Forestry), and Punta Curiñanco Reserve (80ha, administered by the non-governmental organization CODEFF) (Fig. 1). Each of these rural communities have less than 400 inhabitants (INE, 2017) where the main subsistence economic activities are cattle and poultry farming, forestry, artisanal fishing and tourism (CONAF, 2014a).
 Pilolcura, (B) Curiñanco, (C) Bonifacio, (D) Chaihuín, (E) Huiro and (F) Cadillal. Map shows wooded areas (native forests and plantations) in green and other land uses in grey. Note: Only the northernmost sections of the Valdivian Coastal Reserve and Alerce Costero National Park are shown.")
Study area including six rural communities adjacent to protected areas in the coastal range of Los Ros region, Chile; (A) Pilolcura, (B) Curiñanco, (C) Bonifacio, (D) Chaihuín, (E) Huiro and (F) Cadillal. Map shows wooded areas (native forests and plantations) in green and other land uses in grey. Note: Only the northernmost sections of the Valdivian Coastal Reserve and Alerce Costero National Park are shown.
Between April, 2016 and July, 2017 we surveyed households that reported having at least one domestic cat (n=67), to characterize the cat population in the study area. Households were chosen through an opportunistic sampling method, where houses located closer to protected areas were sampled first, to then continue sampling houses located increasingly further away from forest and wooded areas. The main constraints were the absence of household residents at the time of sampling and willingness of cat owners to participate in this study.
For each household we investigated the average number of cats (and other pets) per house, and the geographic origin of each cat to evaluate pet movement between locations and other demographic data. We also assessed care provided by the owners; health management and treatments, birth control and type of food provided. We recorded the roles the owners assigned to the cats and asked owners their opinion towards neutering. To obtain this information we used a set of open-ended questions based on a reference questionnaire designed for previous studies conducted in the area (Silva-Rodríguez and Sieving, 2011; Villatoro et al., 2016).
GPS tracking of domestic cats from rural communitiesBetween April, 2016 and July, 2017 we fitted GPS CatLog® Gen 2 tracking devices (Catnip Technologies, USA) to 68 owned, free-roaming domestic cats in the study area. These lightweight low-cost data loggers are specially designed for this species and have been described to have high fix success rates and position accuracy, supporting their use in studies tracking free-ranging animals (Forin-Wiart et al., 2015; Morris and Conner, 2017). Devices were positioned on the cats’ backs using a soft fabric harness with Velcro straps, which prioritized their safety. Devices were configured, based on Bengsen et al. (2012), to record one fix per hour, 24hours a day, so that battery life could last a minimum of 30 continuous days. We tracked between 5 and 18 cats per locality, the inclusion criteria being obtaining a signed agreement from the owners to participate in the study (through an Informed Owner Consent Form), the size of the cat for an adequate fitting of the harness and the ability to capture the animal. The procedure of placing GPS devices was accomplished through physical restraint (n=66) when possible; less manageable cats were captured with Tomahawk traps and anesthetized with a combination of xylazine (1mg/kg) and ketamine (10–20mg/kg) (n=2). All procedures were conducted under considerations of animal welfare and ethical aspects and with the approval of the Animal Ethics committee of the Institute of Ecology and Biodiversity in Universidad de Chile, resolution of 20 November 2015.
Upon completion of the study and after the retrieval of each GPS device, we applied a short post-study questionnaire to cat owners to assess any perceived changes in their cats’ behavior while using the harness and device. We aimed to identify possible biases due to reduced movement behavior or nuisances related to the use of the harness.
Domestic cat spatial data analysisWe truncated locations recorded after the date on which the GPS device was recovered in the field or, alternatively, on the date on which the GPS device was removed by the cat or owner, to avoid including locations outside the effective tracking period. We filtered 2-D locations with dilution of position (PDOP) value >5, an approach recommended by Lewis et al. (2007) and supported by other studies (Recio et al., 2011), which aims to reduce location error within the dataset while minimizing the loss of accurate locations.
We identified the land cover at which the GPS fixes were obtained as native forest, plantations, shrubland or grassland and villages, to assess the habitat use of free-roaming domestic cats, using a public database (CONAF, 2014b) in ARCGIS (ver.10.3) software. Land use information contained in this database dates to 2013, with a minimum spatial resolution of 0.5 hectares for forested areas, while areas adjacent to National Parks are mapped with a spatial resolution of 1ha. To investigate further the use of wooded areas by domestic cats, we obtained the mean availability of wooded areas by calculating the average proportion of forest or plantations contained in the estimated home range area for each cat, and mean occupancy, corresponding to the percentage of tracking locations recorded within wooded areas.
We estimated the home range area for each individual domestic cat using 95% fixed Kernel density estimators (Worton, 1989) using the adehabitatHR package (Calenge, 2015) for R (R Core Team, 2019). This method has been widely used (Laver and Kelly, 2008), as it provides more precise estimations compared to other methods (Seaman and Powell, 1996) and it has been recommended for consistency among studies of home range areas (Laver and Kelly, 2008). We used the ad hoc method for smoothing parameter selection. Autocorrelation between locations was addressed by determining a sampling frequency that could better represent the relative amount of time spent in different areas throughout the day, while maximizing the number of tracking days to get more accurate estimates (Fieberg, 2007). Home range estimates were calculated only for those cats with 15 or more days of monitoring, considering evidence from feral cats in which the asymptote in the accumulated area curve was reached between three and 18 days (Recio et al., 2010), and consistent with the conclusions of Leo et al. (2016), who recommended a minimum of 14 days of monitoring. It was also of our interest to assess foraying behaviors and use of wooded areas (as in Sepúlveda et al., 2015), for which we calculated the maximum distance from the household, maximum penetration distance into wooded areas (native forests and plantations) and standard distance using a one standard deviation radius, which provides a distance value that summarizes the dispersion of locations and percentage of locations in wooded areas.
Factors associated with spatial behavior of domestic catsWe used linear mixed effect models (LMM) to explore associations between four response variables describing the spatial extent and land use patterns of domestic cats, including: (i) home range area (ha); (ii) maximum foray distances (m); (iii) maximum distance into forest (m) and (iv) percentage of locations in forest, and six predictor variables including: (i) sex; (ii) age; and (iii) reproductive status (neutered vs. entire) of cats; (iv) type of food provided (mixed vs. commercial only); (v) house density and (vi) distance from house to forest edge (m). Only cats with an effective tracking period of 15 days or more were included for more accurate results. Cat-level predictor variables were obtained through interviews. House density was obtained by counting roofs within a circular area of 100m radius centered at each household based on satellite images of the World Imagery basemap (Esri, , 2019), coupled with field observations to increase accuracy. The area of 100 meters of radius was determined based on the primary area used by most cats in the study and median home ranges observed.
LMMs were fitted using the lme4 package (v. 1.1-21; Bates et al., 2015) in R (v. 3.5.3) programming environment (R Core Team, 2019). To test potential effects of sampling effort on the estimation of response variables, we included alternatively the number of tracking days or the number of tracking locations (both log-transformed) as fixed effects in the fitted LMM. The rural community or ‘locality’ was included as a random factor to account for spatial aggregation of data and potential effects of unrecorded local factors. Considering that most cats belonged to different households (n=21), we did not include ‘household’ (nested within locality) as an additional grouping factor.
We used a simplified backward stepwise model selection algorithm. First, we fitted two alternative full models and performed residual analyses, including either the number of tracking days or the number of tracking locations to account for sampling effort. In most cases response variables were log-transformed to improve homogeneity of variance and distribution of residuals, except for percentage of locations in wooded areas, which was arcsine-square root transformed (Figs. S1 to S5 in Supplementary Material). To correct non-linearity in the residuals (Fig. S5 and Table S1 in Supplementary Material), distance from household to forest edge was log-transformed in models estimating the variance of the percentage of locations in forest. The best full model was selected based on residual analysis, coefficient of determination (R2), and/or Akaike's Information Criterion corrected for small sample size (AICc), using the AICcmodavg R package (v. 2.2-1; Mazerolle, 2019). Then we removed one non-significant parameter at a time from the selected models, based on decreasing p-values. After obtaining a simplified model including only significant parameters (i.e. two or three), all potential nested models were fitted, including the null model (i.e. model including a general intercept and the random factor only) (Table S1 in Supplementary Material). We estimated the marginal and conditional coefficients of determination (R2M and R2C, respectively) for model selection, and assessed both using the explanatory power of fixed effects and the extent of heterogeneity in the response variables among rural communities (i.e. the effect of the random effect), using the rsq (v. 2.0) R package (Zhang, 2020).
Camera trap monitoring data from adjacent protected areasAs a complementary approach to cat tracking, we used the 2016 data of the camera trap monitoring program of the Alerce Costero National Park and Valdivian Coastal Reserve to determine whether cats entered protected areas. The design is available in Silva-Rodríguez et al. (2015, 2018). In 2016, 57 cameras operated in the Valdivian Coastal Reserve and 20 in Alerce Costero National Park (Silva-Rodríguez et al., 2018). Following the monitoring plan, data were truncated after 30 days. Therefore, the sampling effort that we report in this study is 2,257 camera days. Here we report the proportion of camera traps that detected domestic cats. For each camera where cats were detected, we also report other species that could be potentially vulnerable to cats, including prey species and wild felids such as the guigna. For each guigna camera trap record we generated a circular area with a radius of 1km, based on the average home range estimated for this species by Eguren (2012) in the same study area. We used these buffer areas to explore the possibility of spatial overlap with domestic cat tracking data by obtaining the number and frequency of domestic cat fixes within guigna buffers.
ResultsCharacterization of local domestic cat populationWe surveyed a total of 67 households, ranging from 3 to 28 per locality, recording a total of 168 cats (Table 1), with an average of 2.6 cats per house (ranging from one to seven). Ninety-one (54.2%) of the cats were female; the mean age of the cats was two years, ranging between two months and 13 years.
Surveyed houses and cats successfully monitored with GPS tracking devices.
| Total number of housesa | Surveyed houses | Total number of cats in surveyed houses | Cats monitored with GPS | |
|---|---|---|---|---|
| Chaihuín | 94 | 13 (13.8%) | 40 | 18 (45.0%) |
| Huiro | 37 | 28 (75.7%) | 53 | 6 (11.3%) |
| Cadillal | 16 | 8 (50.0%) | 19 | 5 (26.3%) |
| Curiñanco | 150 | 3 (2.0%) | 8 | 5 (62.5%) |
| Pilolcura | 13 | 11 (84.6%) | 34 | 9 (26.5%) |
| Bonifacio | 47 | 4 (8.5%) | 14 | 5 (35.7%) |
The geographic origin of the cats was mainly local (89.3%); most were born in the same or adjacent localities and only 10.7% were brought from other cities or towns. Participants reported that during the year before the interview 71 kittens were born. The average number of kittens per litter was three (range: 1–5) and the average number of births per female was 0.7 (range: 0–4). The total number of cats that died during the previous year was 17, and in addition, 26 cats were lost.
Most of the owners assigned a dual role to their cats; as companion animals and for rodent control (63.1%). Health care and management were generally deficient, as a high proportion of the cats were lacking both vaccinations and antiparasite treatments (85.7%). Regarding feeding, 72.3% of cats received a mix of low-quality commercial cat dry food (pellets) and food scraps, 23.2% only commercial cat dry food, 3% only food scraps, while 1.5% received concentrate or cattle food. Birth control was infrequent; only 13.1% of cats were neutered or spayed (18% of female cats, 6.7% of male cats), although 76.1% of the owners were in favor of neutering all their pets (cats and dogs), and 23.9% of them were in favor of neutering only females.
Spatial movements of rural domestic catsWe recovered 53 of the tracking devices fitted to 68 cats; the 15 remaining were lost. We were able to track and obtain useful data from 48 domestic cats (Table 1), with a mean tracking period of 19.2 days (5–55 days). Of the total cats monitored, 22 were females (45.9%) and 26 males (54.2%), 26 were juveniles (54.21%) and 22 adults (45.8%); 21 were monitored during spring-summer (September and December) (43.9%), and 27 during autumn-winter (April, July and August) (56.2%). Tracked cats belonged to 35 different houses. In 68.6% of cases we monitored a single cat, in 28.6% we monitored two, and in only 2.9% we monitored three or more cats per household.
The post-study questionnaire to assess possible behavioral changes associated with the use of the harness and device resulted in 64.3% of cat owners claiming that the device did not affect their cat's normal behavior, while 70.3% reported that it did not bother the cat in a significant way. In addition, 88.3% affirmed the device did not alter the cat's appetite, 72% said the cat moved normally, and 90.7% that the cat continued using the usual sites.
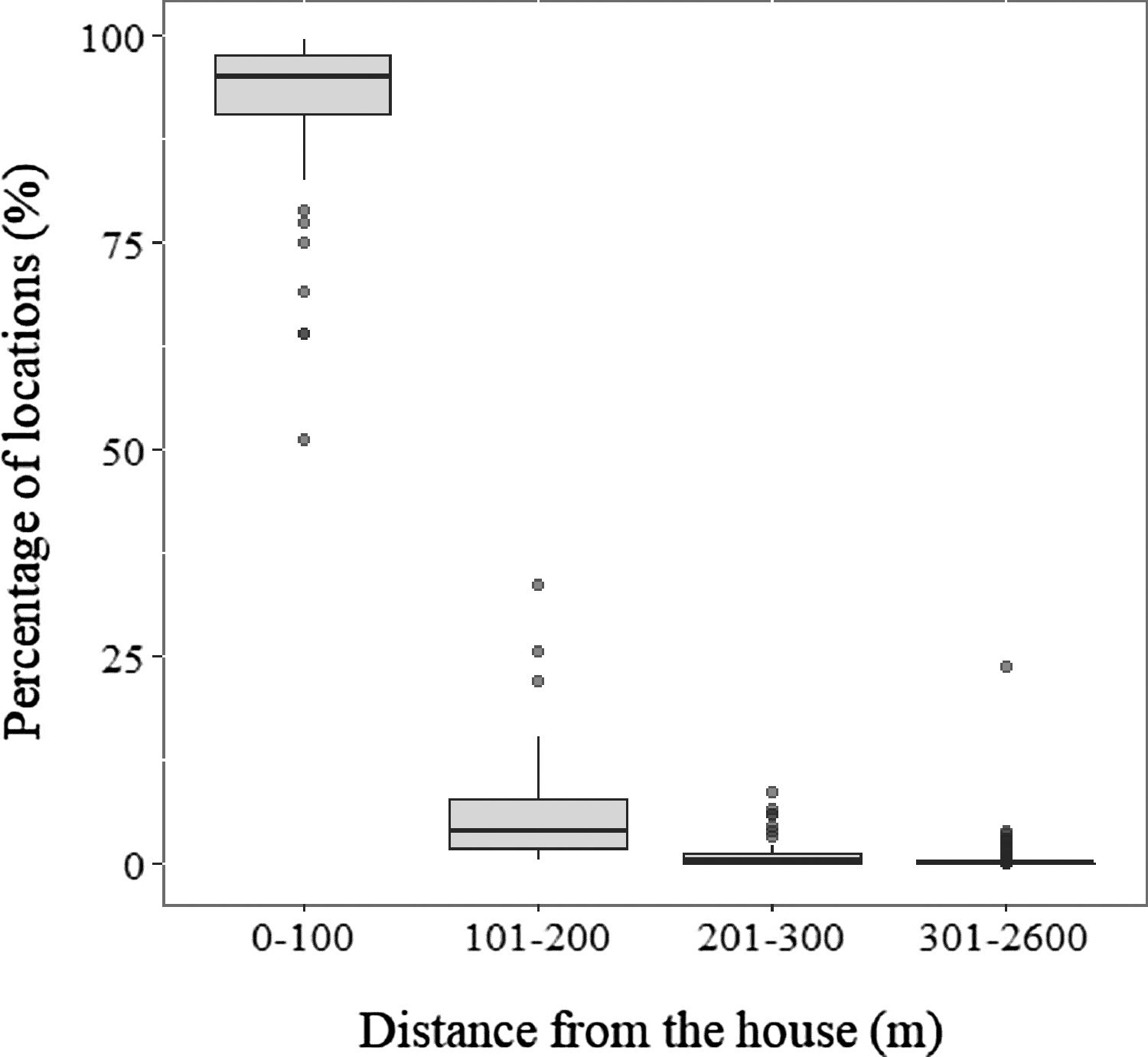
We obtained a total of 19,273 GPS fixes from the 48 monitored domestic cats, with an average of 404.7 (±213.4) per domestic cat. After filtering the GPS locations, we obtained 19,207 fixes that were used for analysis. Of the total fixes, 91% were obtained within 100 meters from the household, considered as their core movement area. As the distance from the household increased, the percentage of fixes for each cat decreased (Fig. 2). Nevertheless, all cats registered fixes further than 100 meters from their house, with one cat ranging as far as 2.5km (Fig. 2, Table 3).
. Most cats spent over 90% of the time close to households, as a larger proportion of locations were recorded within 100m for most cats. Rarely, the proportion of locations near the house was lower (between 50% and 75%), which was usually associated with repeated locations around nearby houses located at distances greater than 100m.")
Percentage of locations per cat obtained at different distance ranges from the house (m). Most cats spent over 90% of the time close to households, as a larger proportion of locations were recorded within 100m for most cats. Rarely, the proportion of locations near the house was lower (between 50% and 75%), which was usually associated with repeated locations around nearby houses located at distances greater than 100m.
Summary statistics of explanatory variables for 35 cats.
| Mean | Median | SD | Min | Max | |
|---|---|---|---|---|---|
| Home range (ha) | 7.2 | 3.3 | 9.8 | 1.0 | 47.2 |
| Maximum distance from house (m) | 738.6 | 565 | 554 | 108 | 2,534 |
| Maximum distance into wooded areas (m) | 269.4 | 160 | 314.5 | 0 | 1,336 |
| Locations in wooded areas (%) | 12.1 | 5.1 | 20.6 | 0 | 97.2 |
| Standard distance (m) | 109.4 | 74.2 | 88 | 36.1 | 372.7 |
Abbreviations: SD: Standard deviation.
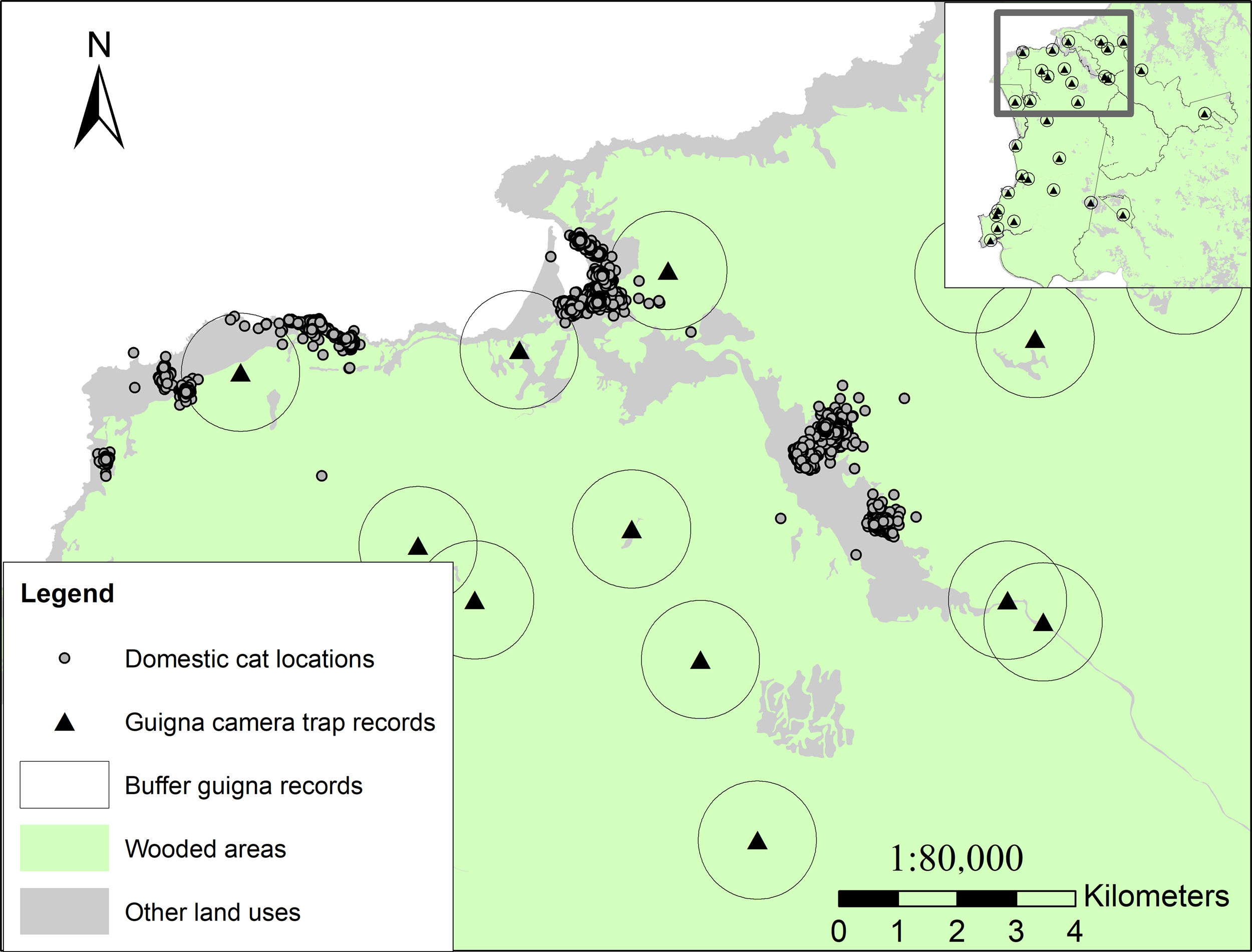
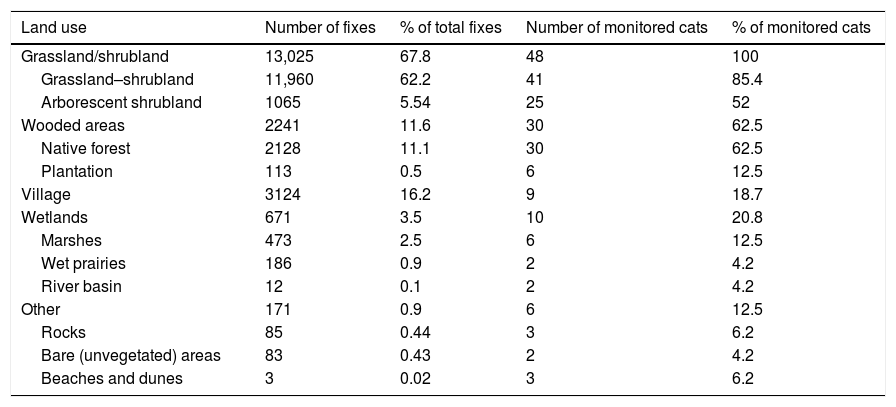
Most of the fixes (67.8%) were obtained in grassland or pasture, corresponding to the land use where most houses are located, and were used by all cats (n=48). Locations obtained in native forest represented 11.1% of the total fixes and were used by 30 of the monitored cats (62.5%). We obtained 16.2% of locations in villages, referred to as areas where different services, businesses and some of the houses are located, and these areas were used by 9 monitored cats (18.7% of fixes). Locations within plantations represented less than 1% of the total fixes and were used by only six of the monitored cats (12.5%). We also recorded locations in wetland areas (3.5%), including marshes and river basins. Bare areas such as beaches and dunes represented less than 1% of the fixes (Table 2). A total of 329 cat locations were recorded inside three protected areas including Alerce Costero National Park, Valdivian Coastal Reserve and Punta Curiñanco Reserve, which accounted for 1.7% of the total fixes, and these areas were used by 9 of the monitored cats (18.7%). We recorded 459 fixes (2.3%) within 1km of locations where guignas were detected through camera trapping, corresponding to locations of 13 of the monitored cats (26.5%) near Alerce Costero National Park and Valdivian Coastal Reserve (Fig. 3). The mean availability of wooded areas within cats’ home ranges was 19.2% (Q1=0%, Q3=33.04%), while the mean occupancy was 9.7% (Q1=0%, Q3=10.6%).
Locations of domestic cats in different land use areas.
| Land use | Number of fixes | % of total fixes | Number of monitored cats | % of monitored cats |
|---|---|---|---|---|
| Grassland/shrubland | 13,025 | 67.8 | 48 | 100 |
| Grassland–shrubland | 11,960 | 62.2 | 41 | 85.4 |
| Arborescent shrubland | 1065 | 5.54 | 25 | 52 |
| Wooded areas | 2241 | 11.6 | 30 | 62.5 |
| Native forest | 2128 | 11.1 | 30 | 62.5 |
| Plantation | 113 | 0.5 | 6 | 12.5 |
| Village | 3124 | 16.2 | 9 | 18.7 |
| Wetlands | 671 | 3.5 | 10 | 20.8 |
| Marshes | 473 | 2.5 | 6 | 12.5 |
| Wet prairies | 186 | 0.9 | 2 | 4.2 |
| River basin | 12 | 0.1 | 2 | 4.2 |
| Other | 171 | 0.9 | 6 | 12.5 |
| Rocks | 85 | 0.44 | 3 | 6.2 |
| Bare (unvegetated) areas | 83 | 0.43 | 2 | 4.2 |
| Beaches and dunes | 3 | 0.02 | 3 | 6.2 |
 camera trap records with 1km buffer areas within Alerce Costero National Park and Valdivian Coastal Reserve. Map shows wooded areas (native forests and plantations) in green and other land uses in grey. Domestic cat GPS fixes overlap with buffer areas of guigna camera trap records (n=459), indicating proximity of <1km between individuals of these species, mostly in areas close to households and forest edges. Note: map inset shows all guigna camera trap records within the two protected areas.")
Domestic cat GPS fixes and guigna (L. guigna) camera trap records with 1km buffer areas within Alerce Costero National Park and Valdivian Coastal Reserve. Map shows wooded areas (native forests and plantations) in green and other land uses in grey. Domestic cat GPS fixes overlap with buffer areas of guigna camera trap records (n=459), indicating proximity of <1km between individuals of these species, mostly in areas close to households and forest edges. Note: map inset shows all guigna camera trap records within the two protected areas.
Maximum linear distances recorded from the house ranged between 108 and 2,534m, with a median distance of 565m (Q1=343m, Q3=977m). Maximum penetration distance into wooded areas (native forest and plantations) ranged from 0 to 1,336m, with a median of 160m (Q1=37m, Q3=407). Domestic cat home range areas varied between 1.0 and 47.2 hectares, with a median of 3.3ha (Q1=2.3ha, Q3=5ha) (Table 3).
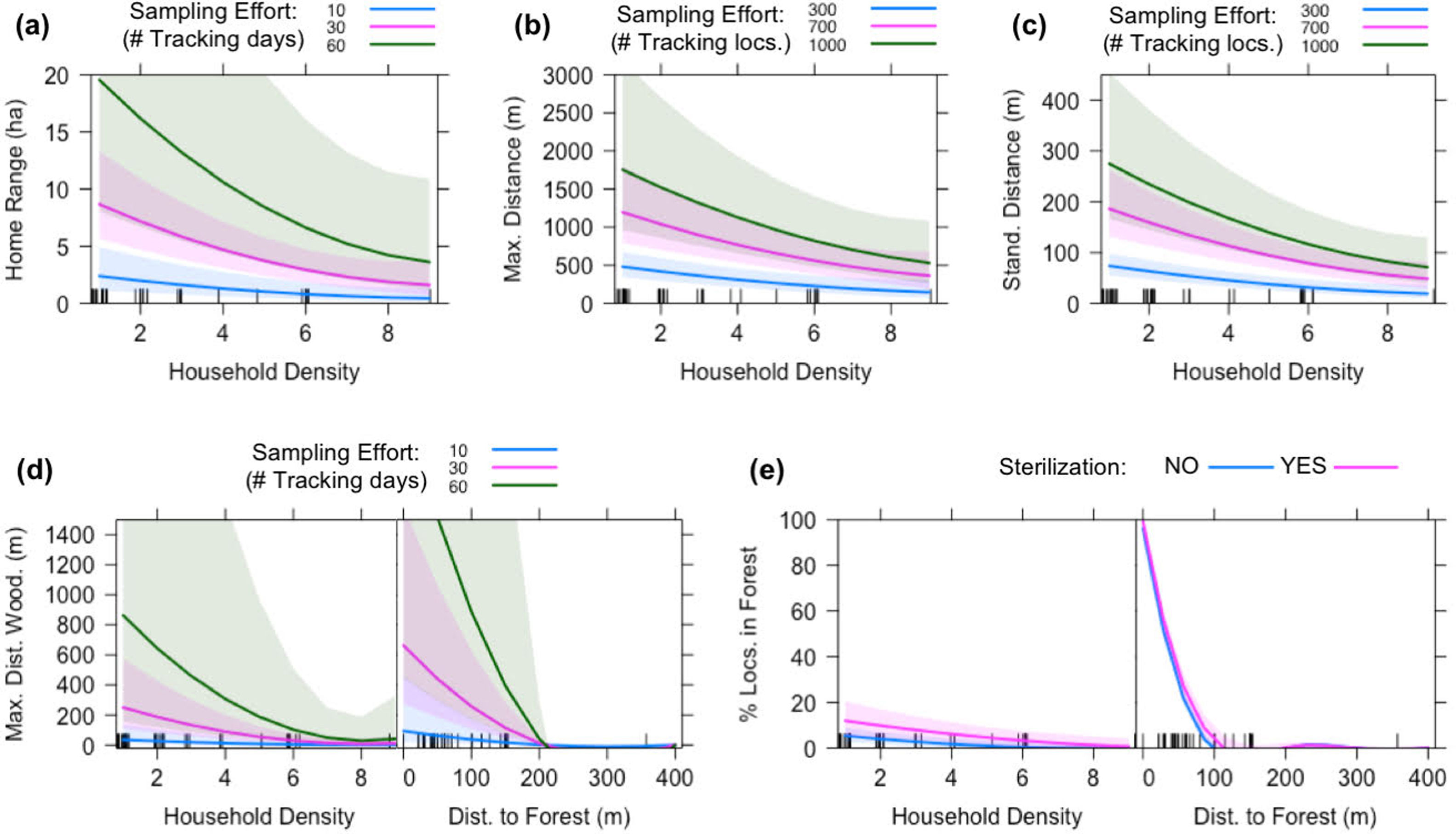
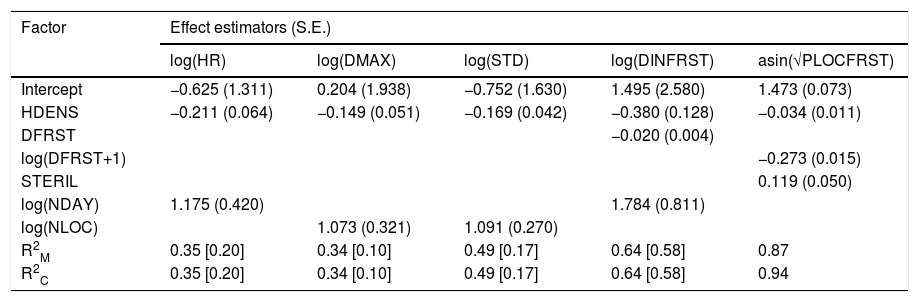
Factors associated with spatial behavior of domestic catsTen to seventeen linear mixed effect models were fitted for each of the response variables associated with cat movements (see Table S1 in Supplementary Material). The selected models show that all variables responded negatively to house density (Fig. 4, Table 4). There was a strong and negative association with the distance of the house to forest edge for the two variables associated with the use of wooded areas, including the maximum penetration distance and the percentage of locations, which dropped to almost zero when houses were located at distances greater than 200m and 100m, respectively (Fig. 4d-e, Table 4). Reproductive status had a subtle effect on the percentage of locations in wooded areas, with neutered individuals registering more locations in these areas (Fig. 4e, Table 4).
. The tick marks on the X axis show the distribution of the data. House density had a significant negative effect on all response variables, including home range, maximum distance, standard distance, maximum distance into wooded areas and percentage of locations in wooded areas. The distance of the house to the forest edge showed a strong negative association with maximum distance into wooded areas and percentage of GPS fixes in wooded areas.")
Partial effect plots for the selected linear mixed effect models. The Y axis is the partial effect of the predictor variable on the response variables (X axis). The tick marks on the X axis show the distribution of the data. House density had a significant negative effect on all response variables, including home range, maximum distance, standard distance, maximum distance into wooded areas and percentage of locations in wooded areas. The distance of the house to the forest edge showed a strong negative association with maximum distance into wooded areas and percentage of GPS fixes in wooded areas.
Fixed effect factors included in the selected linear mixed effect models (LMM) for each response variable.
| Factor | Effect estimators (S.E.) | ||||
|---|---|---|---|---|---|
| log(HR) | log(DMAX) | log(STD) | log(DINFRST) | asin(√PLOCFRST) | |
| Intercept | −0.625 (1.311) | 0.204 (1.938) | −0.752 (1.630) | 1.495 (2.580) | 1.473 (0.073) |
| HDENS | −0.211 (0.064) | −0.149 (0.051) | −0.169 (0.042) | −0.380 (0.128) | −0.034 (0.011) |
| DFRST | −0.020 (0.004) | ||||
| log(DFRST+1) | −0.273 (0.015) | ||||
| STERIL | 0.119 (0.050) | ||||
| log(NDAY) | 1.175 (0.420) | 1.784 (0.811) | |||
| log(NLOC) | 1.073 (0.321) | 1.091 (0.270) | |||
| R2M | 0.35 [0.20] | 0.34 [0.10] | 0.49 [0.17] | 0.64 [0.58] | 0.87 |
| R2C | 0.35 [0.20] | 0.34 [0.10] | 0.49 [0.17] | 0.64 [0.58] | 0.94 |
Abbreviations: HR: home range size, DMAX: maximum location distance, STD: standard location distance, DINFRST: distance of penetration into forest, PLOCFRST: percentage of locations recorded within forest, HDENS: house density, DFRST: house-forest distance, STERIL: sterilization NDAY: number of tracking days, NLOC: number of tracking locations. R2M: marginal coefficient of determination (i.e. fixed effects), R2C: conditional coefficient of determination (i.e. fixed+random effects). The coefficients of determination without the effect of sampling effort are shown between brackets.
The selected models accounted for less than half of the observed variability in home range size, maximum and standard distance from the household, and for 20% or less of the variability when sampling effort was not considered (Table 4). Accordingly, the inclusion of sample size was necessary in our modelling framework as these variables responded positively to either the number of tracking days or tracking locations and did not reach an asymptote within the observed range (Figs. 4a–c, Table 4). Conversely, most of the variability observed for the maximum penetration distance and percentage of localization in wooded areas was explained by the selected models, and sampling effort had little or no effect on their estimation (Table 4). Note that spatial behavior of cats was highly consistent across rural communities, as conditional and marginal coefficients of determinations show little or no difference (Table 4).
Camera trap records and additional evidenceWe obtained cat records in only one (1%) of the camera traps, which was located inside the Valdivian Coastal Reserve. In addition, one cat was detected in a different camera trap after the end of the 30-day period of camera trap monitoring in the Alerce Costero National Park. Both cameras that detected cats were relatively close to households. Interestingly, we detected guignas in 40% of the cameras within the 30-day period, including both cameras that detected cats. In one camera, the time difference between records of a cat and a guigna was only one hour (Fig. 5). We also detected rodents and birds with these cameras, including endemic species such as chucao (Scelorchilus rubecula, Fig. 5), black-throated huet-huet (Pteroptochos tarnii), Austral thrush (Turdus falcklandii) and striped woodpecker (Veniliornis lignarius).
 chucao tapaculo (Scelorchilus rubecula) is shown in a site where cats roam (B, C). Cat (left) and guigna (right) recorded in the same location with one hour (Valdivian Coastal Reserve; C, D) and five days difference (Alerce Costero National Park; E, F).")
Camera trap records of domestic cats and wild animals. (A) chucao tapaculo (Scelorchilus rubecula) is shown in a site where cats roam (B, C). Cat (left) and guigna (right) recorded in the same location with one hour (Valdivian Coastal Reserve; C, D) and five days difference (Alerce Costero National Park; E, F).
We incidentally captured three domestic cats while aiming to capture guignas in the aforementioned protected areas; one of them was recaptured five times on different days. The traps were located near roads, trails and the park ranger's cabin. The captured cats were not feral; we were able to identify their homes and owners for at least two of them. Distance of these capture sites to the households ranged from 856 to 1,145m. Guigna camera trap records were also obtained in these areas during the same year (not shown).
DiscussionMost cats in our study had small home ranges, similar to those recorded in other areas of the world (e.g. Barrat, 1997; Metsers et al., 2010; Van Heezik et al., 2010; Kays et al., 2020), but these could be underestimated considering the marked effect of sampling effort in our models. As in other studies (Kays et al., 2020), cats spent most of their time (mean of 90% per cat) within 100 meters of the house. Camera traps deployed in two protected areas detected cats infrequently – far less than guignas – and the few cats captured in these protected areas were not feral. Therefore, our data show that cats concentrate their activity in human-inhabited areas and that their presence in natural areas is mostly limited to forest edges.
Home range size was negatively correlated with house density, which has been identified as an important predictive factor (Hall et al., 2016), likely explained by the fact that cat home ranges are smaller at higher cat densities (Van Heezik et al., 2010; Bengsen et al., 2016). Even though cats concentrate their activity near houses and in anthropized areas (Ferreira et al., 2011; Morin et al., 2018; Kays et al., 2020; this study), this does not prevent them from entering wildlife habitat. We observed that 63% of the cats used wooded areas (mainly native forest) at least occasionally (12% of the fixes on average), and detected foray distances up to 1.3km into native forest. However, the time cats spent in the forest (percentage of locations in forest) rapidly decreased as the distance between the house and forest increased. Our data suggest that potential interactions between forest vertebrates are more likely to occur in the forest edge and in patches located near houses. For example, guignas make intensive use of forest edges, do not appear to avoid houses (Schüttler et al., 2017) or dogs (Moreira-Arce et al., 2015; Gálvez et al., 2018) and may prey on poultry (e.g. Zorondo-Rodríguez et al., 2014). Therefore, guignas may not be avoiding areas used by domestic cats, and domestic cats may roam into guigna habitat, facilitating co-occurrence of these species, as reported here and in other studies (Silva-Rodríguez et al., 2020). This proximity may facilitate interactions and disease transmission from rural domestic cats to guignas, as suggested by previous studies (Mora et al., 2015; Sacristán et al., 2021; Napolitano et al., 2015).
Cat movement was not associated with the variables explored to assess the effect of human care (food provision and neutering), similar to findings from previous studies (Hall et al., 2016). The lack of association between reproductive status and cat home range could be related to the small number of neutered cats included in this study. Therefore, we acknowledge that the effect of neutering on cat movement remains unclear, with some studies suggesting that intact cats have larger home ranges (e.g. Castañeda et al., 2019; Kays et al., 2020) and others suggesting no effects (e.g. Hall et al., 2016; this study). However, we found that neutered cats used forest slightly more often than entire cats (see Fig. 4), but the interpretation of this association requires caution considering the small number of neutered cats we tracked. As stated by Kays et al. (2020), clarifying the potential effects of neutering on cat movement remains as an important research priority.
Our results provide relatively good news for large natural areas far from human settlements, such as protected areas, where the impacts of cats are likely to be marginal, as we did not observe preference of use of wooded areas by domestic cats and cat locations were mainly restricted to forest edges and associated with houses located nearby (less than 200m). Nonetheless, free-roaming domestic cats have a strong potential to emerge as an important threat in rural areas of southern Chile in the upcoming years. Rural areas of southern Chile are facing rapid processes of land subdivision (e.g. Díaz et al., 2010) associated with different factors, among them the development of residential areas (Petitpas et al., 2017; Gálvez et al., 2018). Land subdivision can be negative for wild carnivores such as the guigna (Gálvez et al., 2018) and, as our results suggest, may also increase the exposure of this and other wild species to cats’ impacts. In rural areas, cats are often owned as companion animals and to control rodents (Silva-Rodríguez and Sieving, 2011; this study), therefore it is predictable that if houses are built, cats will be present. Chilean regulations allow rural lands to be subdivided down to 0.5ha (Márquez, 1999), below the size of home ranges reported for owned cats in this and other studies (e.g. Horn et al., 2010; Hall et al., 2015; Kays et al., 2020). Therefore, under ongoing land subdivision processes for residential purposes, high cat densities may be expected. This has important implications. Cats kill less prey than wild felids on a per-capita basis, but predation is concentrated in small areas, making their per-capita impact per area (prey/ha/cat) comparable to those of wild felids (Kays et al., 2020). With land subdivision, cat densities – and their cumulative impacts – will likely be much higher than those of wild carnivores (as reported by Kays et al., 2020).
At a broader scale, the threat that domestic cats pose for native wildlife should be considered in land planning processes near priority conservation areas and habitat of threatened wildlife in human-dominated landscapes. Our results are clear regarding the strong effect of the distance from the forest as a strong predictor of cats’ use of these areas, as we observed that when houses are located at distances greater than 200m from the forest the percentage of cat GPS fixes within these areas dropped to almost zero, similar to what we observed in maximum distances into wooded areas, which dropped to zero when houses were located further than 100m from the forest edge. These results support some landscape management recommendations to reduce cats’ presence and impacts to local wildlife, such as the creation of cat-exclusion zones around reserves (Metsers et al., 2010), which in our study area should be of at least 2.5km, based on the maximum distance recorded. Nevertheless, as stated before, these results could be underestimated, and it is likely that greater distances could be necessary to completely avoid cats’ presence. Considering that cat exclusion zones are likely to be unfeasible in most cases, alternatively, we recommend that cat population management should be prioritized within 2.5km from protected areas.
Locally, movement restriction of owned cats appears as the most important strategy to reduce contact with wild species (Hall et al., 2016). Unfortunately, such management is almost non-existent in rural areas of Chile (Silva-Rodríguez and Sieving, 2011; this study), and – as shown by a study in New Zealand – unlikely to be widely adopted by cat owners (Linklater et al., 2019). Alternatively, strategies should focus on implementing landscape management strategies to reduce cats’ presence and mitigate their impacts in important conservation areas, by reducing fragmentation of remaining natural areas, and managing cat populations around protected areas. Improving care provided by owners can also help to mitigate specific threats such as predation and disease transmission. The risk of exposure to some of the infectious diseases that have been seen to affect native wildlife in Chile, such as retroviruses infecting guignas (Mora et al., 2015; Sacristán et al., 2021), can be managed through vaccination (Horzinek et al., 2013), but this is not an option for reducing the risk of wildlife exposure to other pathogens associated to domestic cats, such as toxoplasmosis, which can cause mortality and negatively impact wild species populations (Miller et al., 2020). To manage predation, the use of collar-mounted devices – as a strategy to reduce predation on birds, mammals and herpetofauna – has shown some positive results (Calver et al., 2007; Calver and Thomas, 2011; Hall et al., 2015), However, evidence on the effectiveness of these and other devices is still limited (Williams et al., 2018) and acceptability by owners should be assessed before promoting them as part of mitigation strategies. The complexity of this issue requires that more research on innovative strategies to reduce the risk of cats to wildlife is conducted. Finally, we caution that the adoption of strategies to reduce cat impacts on wildlife may be challenging, because cat predation – especially on small mammals –may be deemed as positive by many owners (Crowley et al., 2019). Acknowledging the role of cats in rural households and providing effective and sustainable alternatives to rodent control is important to achieve changes in cat management. Finally, reproductive control of the cat population is a fundamental action to reduce and control the number of free-roaming cats. Our results show that most owners are in favor of neutering cats, contrasting with the low proportion of animals neutered/spayed. This points out the need for improving access, in terms of cost and proximity, to these veterinary services in rural areas located within conservation priority regions or threatened species habitat.
ConclusionOur study suggests that even though domestic cats in rural areas of southern Chile rarely roam large distances, they often use native forest located close to their houses. These forests provide habitat for native and endemic wildlife susceptible to cats’ impacts. Therefore, our data suggest that under the ongoing processes of urbanization and land subdivision in rural landscapes of southern Chile, cats could become an emergent threat for native wildlife.
Our results emphasize the need to adopt multiple approaches to control domestic cat populations and mitigate their impacts on wildlife. For example, land planning for residential purposes and for the creation of conservation reserves should consider minimum distances of households from conservation priority areas and threatened species habitat. Regarding management practices to be promoted among cat owners, strategies to limit or reduce ranging behavior are the most effective in reducing cats’ impacts, but implementation at the broader scale is unlikely given their role as pest controllers in rural settings, as described in this study, and negative attitudes of owners towards these strategies (Crowley et al., 2019; Linklater et al., 2019). In rural towns or settlements located near conservation priority areas, immediate actions need to be implemented, such as continuous long-term neutering and vaccination programs to control cat population growth and reduce the prevalence and spread of infectious diseases, mitigating threats to local wildlife.
FundingOur work was funded by CONICYT FONDECYT Iniciación [11150934] (CN), Morris Animal Foundation [D15ZO-413] (CN), National Geographic Society [C309-15] (CN), Mohamed bin Zayed Species Conservation Fund [152510351] (CN), ANID PAI [77190064] (CN), the Wild Felid Association (IS). E.A.S., thanks funding from CONICYT FONDECYT [11171006]. AF thanks CONICYT PIA/BASAL FB0002. MJL thanks ANID Scholarship Program MAGISTER BECAS CHILE/2018–73190501.
We gratefully acknowledge local people that agreed to participate in this study and kindly gave us the opportunity to track their domestic cats. We thank The Nature Conservancy and CONAF for allowing us to use their camera trap monitoring datasets and for logistic support. We thank managers and park rangers of the Valdivian Coastal Reserve and Alerce Costero National Park: Patricia Barría, Liliana Pezoa, Erwin Ovando, Danilo González, Omar Ponce, Rene Cárdenas, who have conducted the field work of the monitoring programs and collaborated with us in the field. We also thank Viviana Vásquez and Brayan Zambrano, who classified camera trap pictures.
The following are the supplementary data to this article:

